Abstract
Human parainfluenza virus type 1 (hPIV1) has two spike glycoproteins, the hemagglutinin-neuraminidase (HN) glycoprotein as a receptor-binding protein and the fusion (F) glycoprotein as a membrane-fusion protein. The F glycoprotein mediates both membrane fusion between the virus and cell and membrane fusion between cells, called syncytium formation. Wild-type C35 strain (WT) of hPIV1 shows little syncytium formation of infected cells during virus growth. In the present study, we isolated a variant virus (Vr) from the WT that showed enhanced syncytium formation of infected cells by using our previously established hPIV1 plaque formation assay. Vr formed a larger focus and showed increased virus growth compared with WT. Sequence analysis of the spike glycoprotein genes showed that the Vr had a single amino acid substitution of Ile to Val at position 131 in the fusion peptide region of the F glycoprotein without any substitutions of the HN glycoprotein. The Vr F glycoprotein showed enhanced syncytium formation in F and HN glycoprotein-expressing cells. Additionally, expression of the Vr F glycoprotein increased the focus area of the WT-infected cells. The single amino acid substitution at position 131 in the F glycoprotein of hPIV1 gives hPIV1 abilities to enhance syncytium formation and increase cell-to-cell spread. The present study supports the possibility that hPIV1 acquires increased virus growth in vitro from promotion of direct cell-to-cell transmission by syncytium formation.
INTRODUCTION
Human parainfluenza virus type 1 (hPIV1) is a respiratory pathogen that causes bronchiolitis, pneumonia, laryngotracheobronchitis and croup in infants or young children.1–4) hPIV1 infections may also cause more severe respiratory diseases, especially in the elderly and immunocompromised patients. However, there is no clinical therapy or vaccine against hPIV1.5) hPIV1 has two spike glycoproteins, hemagglutinin-neuraminidase (HN) glycoprotein as a receptor-binding protein and fusion (F) glycoprotein as a membrane-fusion protein, on the viral surface.6) hPIV1 binds to terminal sialic acids of glycoconjugates on the host cell surface through the HN glycoprotein.6,7) The F glycoprotein is a common spike protein in paramyxovirus and mediates both membrane fusion between the virus and cell (virus–cell fusion) and membrane fusion between cells (cell–cell fusion).8)
Nascent HN and F glycoproteins are abundantly expressed on the cell surface membrane of paramyxovirus-infected cells. The F glycoprotein on the cell surface membrane induces cell–cell membrane fusion between virus-infected cells and their neighboring non-infected cells, resulting in multinucleated giant cell formation, called syncytium formation.1) Paramyxovirus simian virus 5 spreads infection with cell-to-cell direct transmission by syncytium formation.9) Syncytium formation has been reported to be associated with paramyxovirus growth.8,10) Remarkable syncytium formation can be seen in cells infected with some paramyxoviruses such as Sendai virus (SeV), respiratory syncytial virus, and human parainfluenza virus type 3 (hPIV3).11,12) hPIV1 shows little syncytium formation during virus growth in vitro.8,13)
We previously established a hPIV1 plaque formation assay.14) In the present study, we isolated an hPIV1 variant (Vr) that had acquired enhanced syncytium formation ability compared with wild-type C35 strain (WT) of hPIV1 by our plaque formation assay. The Vr showed increased progeny virus release and formed larger plaques than did WT. The Vr had a single amino acid substitution of Ile to Val at position 131 in the fusion peptide region of the F glycoprotein without any substitutions in the HN glycoprotein. The single amino acid substitution at position 131 of the F glycoprotein enhanced syncytium formation and increased the focus sizes. This study showed that the single amino acid substitution at position 131 in the fusion peptide region of the F glycoprotein of hPIV1 gives hPIV1 abilities to enhance syncytium formation and increase cell-to-cell spread in vitro.
MATERIALS AND METHODS
Cells, Viruses, and AntibodiesLewis lung carcinoma-monkey kidney (LLC-MK2) cells were maintained in Eagle’s minimum essential medium (MEM) supplemented with 5% heat-inactivated fetal bovine serum (FBS). Human epidermoid cancer (Hep-2) cells were maintained in Dulbecco’s modified Eagle’s MEM supplemented with 10% FBS. The WT (C35 strain) of hPIV1 was kindly provided by Allen Portner of St. Jude Children’s Research Hospital. The WT was propagated in LLC-MK2 cells. Rabbit anti-hPIV1 antibody was prepared by immunization with the WT as described previously and was purified on a Protein G HiTap™ Protein G HP (CE Healthcare Life Sciences, Piscataway, NJ, U.S.A.).15)
Isolation of a Variant Virus Showing Obvious Syncytium FormationThe plaque assay of the WT was carried out as in our previous study using confluent monolayers of LLC-MK2 cells.14) A plaque in which obvious syncytium formation was observed under a microscope was selected, and then the Vr was obtained from the plaque and was inoculated into newly confluent monolayers of LLC-MK2 cells in a 6-well plate. The cells were incubated in hybridoma-serum-free medium complete dry powdered medium (SFM, Thermo Fisher Scientific Inc., Waltham, MA, U.S.A.) containing 3 µg/mL of acetylated trypsin at 37°C for 3 d. The culture medium was harvested and was added to newly confluent monolayers of LLC-MK2 cells in a 6-well plate. The cells were incubated in SFM containing 3 µg/mL of acetylated trypsin at 37°C for 3 d again. The plaque assay was again performed with the harvested culture medium. Finally, the Vr showing obvious syncytium formation was isolated from the plaque formed by the WT.
Construction of an Expression Plasmid Vector Containing F or HN GeneF genes and HN genes were cloned into the pCAG GS/MCS expression vector described previously.8) Briefly, each viral RNA was extracted from virus particles with TRIzol reagent (Invitrogen Corp., Carlsbad, CA, U.S.A.) according to the manufacturer’s instructions. Full-length cDNAs of the F gene and HN gene from hPIV1 were amplified by using a TaKaRa RNA PCR™ kit Ver. 3.0 (TaKaRa Bio Inc., Shiga, Japan) with the primers described in Supplementary Table 1. The F genes and HN genes were inserted into the multiple cloning region between the EcoRI site and XhoI site of the pCAG GS/MCS expression plasmid vector. Sequence analysis of the F gene and the HN gene was performed by using ABI Prizm 310 Genetic Analyzer (Life Technologies, Calsbad, CA, U.S.A.) with the primers shown in Supplementary Table 2, and the nucleotide substitution of the Vr was identified.
Focus-Forming Assay and Virus TitrationConfluent monolayers of LLC-MK2 cells in a 12-well plate were washed with phosphate buffered saline (PBS) and were inoculated with serial dilutions of a virus suspension at room temperature for 30 min. The wells were washed with PBS and the cells were overlaid with 2 mL of SFM containing 3 µg/mL acetylated trypsin and 1.2% microcrystalline cellulose Avicel (FMC Corporation, Philadelphia, PA, U.S.A.) at 37°C for 2 d. The cells were fixed with 800 µL of cold methanol for 5 min and reacted with rabbit anti-hPIV1 antibody at room temperature for 30 min and then with horseradish peroxidase-conjugated Protein A (Sigma-Aldrich, St. Louis, MO, U.S.A.) at room temperature for 30 min. Focus-forming units (FFU) as infectious virus titers were calculated from the immunostained foci developed as described previously.16)
A focus-forming assay of WT was also performed in F glycoprotein-expressing cells. Seventy percent confluent monolayers of LLC-MK2 cells in a 12-well plate were transfected with 1 µg of pCAG GS vectors containing the F gene from WT (F-WT gene) or F gene from Vr (F-Vr gene) using the transfection reagent TransIT-LT1 (Mirus, Madison, WI, U.S.A.) according to the manufacturer’s instructions. Cells were also mock-treated without plasmid vectors as a control. After incubation at 37°C for 6 h, the transfected cells were washed with PBS and then the cells were incubated with 1 mL of SFM. After incubation for 22 h, the transfected cells were washed with PBS and the focus-forming assay was performed with WT at a multiplicity of infection (MOI) of 0.0001 for 3 d incubation. The area of the stained focus was measured using Image J release 1.40 g (National Institutes of Health, U.S.A., http://rsb.info.nih.gov/ij/) and represented as the relative focus area to the median of the control focus areas.
Estimation of Multi-Step Growth of the VirusConfluent monolayers of LLC-MK2 cells on a 24-well plate were infected with viruses at a MOI of 0.001 for 30 min at room temperature. After washing the viruses with 250 µL of PBS, the cells were incubated with 500 µL of SFM containing 3 µg/mL of acetylated trypsin. Supernatants of the cells were recovered at 12, 24 and 36 h. Virus titers in the supernatants were measured by a focus-forming assay with triplicated experiments.
Syncytium Formation AssayThe hPIV1 syncytium formation assay (cell-cell fusion assay) was carried out as in our previous study.8) Briefly, for quantification of syncytium formation of hPIV1-infected cells, confluent monolayers of LLC-MK2 cells in a 24-well plate were infected with each virus in SFM at an MOI of 1 at room temperature for 30 min. The plates were incubated at 37°C for 48 h in culture of SFM containing 3 µg/mL of acetylated trypsin until visualization of syncytium formation. The infected cells were fixed with 500 µL of cold methanol for 5 min and then treated with 500 µL of Carrazzi’s haematoxylin solution in PBS (1 : 1, v/v) (Wako Pure Chemical Industries, Ltd., Osaka, Japan) in order to visualize the nuclei. Fusion efficiency (%) of the infected cells was calculated as percentage of the number of nuclei in the syncytia relative to the sum total of nuclei in all cells under 9 optical observations at a magnification of 100.
For quantification of syncytium formation of LLC-MK2 cells expressing both F and HN genes, 70% confluent monolayers of LLC-MK2 cells in a 24-well plate were co-transfected with 1 µg of both pCAG GS vectors containing F and HN genes using the transfection reagent TransIT-LT1 according to the manufacturer’s instructions. After incubation at 37°C for 6 h, the co-transfected cells were washed with PBS and then the cells were incubated with 500 µL of SFM containing 3 µg/mL of acetylated trypsin. After incubation for 46 h, the co-transfected cells were washed with PBS. Fusion efficiency (%) of the co-transfected cells was calculated as percentage of the number of nuclei in the syncytia relative to the sum total of nuclei in all cells under 5 optical observations at a magnification of 100.
For quantification of syncytium formation of Hep-2 cells expressing both F and HN genes, 70% confluent monolayers of Hep-2 cells in a 24-well plate were co-transfected with 500 ng of both pCAG GS vectors containing F and HN genes using the transfection reagent according to the manufacturer’s instructions. After incubation at 37°C for 48 h, the co-transfected cells were treated with 3 µg/mL of acetylated trypsin at 37°C for 30 min. The cells were washed with PBS and incubated with 500 µL of SFM. After incubation for 10 h, the co-transfected cells were washed with PBS. Fusion efficiency (%) of the co-transfected Hep-2 cells was calculated as percentage of the number of nuclei in the syncytia relative to the sum total of nuclei in all cells under 5 optical observations at a magnification of 100.
RESULTS AND DISCUSSION
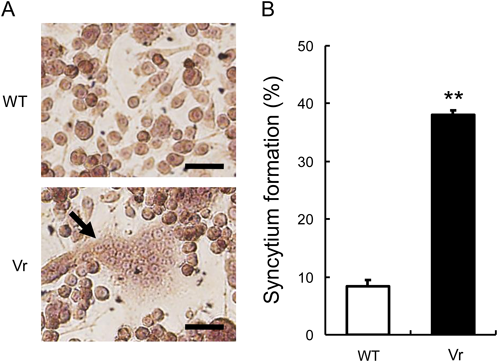
We previously established a hPIV1 plaque formation assay.14) hPIV1 generally shows little syncytium formation in vitro.8,13) However, in our plaque formation assay of hPIV1, there were a few plaques in which obvious syncytium formation was observed (a few syncytium-forming plaques per 100 non-syncytial plaques). It seemed that hPIV1 fortuitously acquired mutations that induced enhanced syncytium formation, by continuously passing the virus in LLC-MK2 cells. We isolated a variant virus (Vr) showing enhanced syncytium formation by selecting a syncytium-forming plaque. To compare the syncytium formation activities of the Vr and the parent C35 strain of hPIV1 (WT), LLC-MK2 cells were infected with each virus at an MOI of 1 and incubated in the presence of acetylated trypsin until clear appearance of syncytia. Whereas syncytium formation was hardly observed in the WT-infected cells, obvious syncytium formation was observed in the Vr-infected cells (Fig. 1A). The Vr showed approximately 4-times higher syncytium formation activity than that of the WT (Fig. 1B).
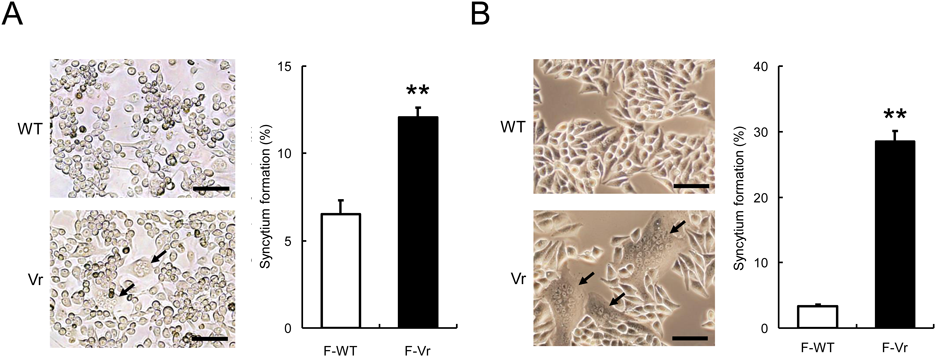
It has been reported that amino acid substitutions of the viral spike HN and F glycoproteins enhance fusion activity of SeV, hPIV1, and hPIV3.8,12,17) We therefore compared amino acid sequences of the HN glycoprotein and the F glycoprotein between the Vr and the WT. The results revealed that the Vr carried a single amino acid substitution of Ile to Val at position 131 in the F glycoprotein without any amino acid substitutions in the HN glycoprotein. According to the amino acid residue alignment with the F glycoprotein from parainfluenza virus type 5 (PIV5) whose three-dimensional structure has been determined,18) the amino acid position 131 of F glycoprotein of hPIV1 was located in the fusion peptide region which is directly inserted into the target membrane for the membrane fusion process. To determine whether the single amino acid substitution in the F glycoprotein of the Vr enhances syncytium formation of hPIV1, the F gene of the WT (F-WT gene) or the Vr (F-Vr gene) was co-transfected together with the HN gene of the WT (HN-WT gene) into LLC-MK2 cells. The transfected cells were incubated in the presence of acetylated trypsin until clear appearance of syncytia. The cells with the HN-WT glycoprotein and the F-WT glycoprotein showed less syncytium formation. On the other hand, the cells with the HN-WT glycoprotein and the F-Vr glycoprotein showed approximately 2-times higher syncytium formation activity than that of the cells with the HN-WT and F-WT glycoproteins (Fig. 2A). Transfection with each F gene alone without the HN gene induced no syncytium formation, indicating that receptor binding activity of the HN glycoprotein is needed for the membrane fusion process of F-WT and F-Vr glycoproteins. For Hep-2 cells, which show obvious syncytium formation in hPIV3 infection, co-transfection with the F-Vr gene together with the HN-WT gene also induced approximately 8-times higher syncytium formation activity than that with HN-WT and F-WT genes (Fig. 2B). These results demonstrated that the amino acid substitution of Ile to Val at position 131 in the hPIV1 F glycoprotein gave enhanced syncytium formation ability in different cell lines, LLC-MK2 and Hep-2 cells. It seemed that expression of HN-WT and F-WT glycoproteins showed less syncytium formation in Hep-2 cells than in LLC-MK2 cells. In our previous study, we used LLC-MK2 cells and Hep-2 cells for establishment of a WT hPIV1 plaque assay.14) Obvious plaque formation is dependent largely on cell death and/or syncytium formation of infected cells. We succeeded in achieving WT plaque formation in LLC-MK2 cells but not in Hep-2 cells. For WT infection, Hep-2 cells showed lower rates of cell death and syncytium formation than did LLC-MK2 cells. It is thought that Hep-2 cells have less fusion efficiency in the case of WT hPIV1 infection.
We compared the amino acid sequences near the position 131 of F glycoprotein among 9 kinds of paramyxoviruses, including the hPIV1 WT and the Vr. The amino acid residue at the position 131 is Ile in hPIV1 WT, human parainfluenza virus type 2, hPIV3, SeV, and Newcastle disease virus, or Val in hPIV1 Vr, human parainfluenza virus type 4, PIV5, and mumps virus. Among these viruses, the fusion peptide region between 113 and 135 is highly conserved19) (Fig. 3). For paramyxoviruses other than hPIV1, the I131V substitution is also suggested to increase fusion activity of each F glycoprotein, which enhances syncytium formation.
It has been reported that enhanced syncytium formation activity promotes multi-step growth of hPIV18) and cell-to-cell spread of Simian virus 5.9) To evaluate virus growth of the Vr, we estimated the focus size as cell-to-cell spread of infection and quantified the supernatant virus titer of infected cells as multi-step growth. Focus sizes of the Vr were significantly larger than those of the WT (Figs. 4A, B). The Vr also significantly increased progeny virus titer in the culture supernatant of infected LLC-MK2 cells during multi-step growth compared to that in the case of the WT (Fig. 4C). These results showed that an enhanced syncytium-forming hPIV1 variant, Vr, increased cell-to-cell spread and multi-step growth in vitro.
To examine whether the single amino acid substitution of Ile to Val at position 131 in the F-Vr glycoprotein affects cell-to-cell spread of infection, we measured focus sizes when F-Vr glycoprotein-expressing cells were infected with the WT. The promotive effect of syncytium formation on direct cell-to-cell transmission of hPIV1 should result in larger focus sizes. LLC-MK2 cells were transfected with the F-WT gene or the F-Vr gene and then infected with the WT. After culture for 3 d, focus sizes were compared. Both F-WT glycoprotein expression and F-Vr glycoprotein expression increased focus areas of the WT compared to that with no transfection of F gene. F-Vr glycoprotein expression resulted in a statistically significant enlargement of focus areas of the WT compared to that with F-WT glycoprotein expression (Fig. 5). The results showed that a single amino acid substitution of Ile to Val at position 131 in the hPIV1 F glycoprotein increased cell-to-cell spread of hPIV1 in vitro.
CONCLUSION
In the present study, we isolated a Vr showing enhanced syncytium formation ability and increased virus growth in vitro by using our established hPIV1 plaque formation assay. The Vr is thought to have appeared by natural occurrence during several passages of the WT. We identified the single amino acid substitution of Ile to Val at position 131 of the fusion peptide region in the F glycoprotein carried by the Vr. The single amino acid substitution of the F glycoprotein gave the hPIV1 abilities to enhance syncytium formation and to increase cell-to-cell spread of virus infection in vitro. The present study supports the possibility that hPIV1 acquires increased virus growth from promotion of direct cell-to-cell transmission by syncytium formation.
Acknowledgments
Keijo Fukushima was a recipient of a scholarship by Honjo International Scholarship Foundation. This work was supported by JSPS KAKENHI, Series of single-year Grants, 2510690, in part, and by THE ICHIRO KANEHARA FOUNDATION, a JSPS KAKENHI Grant (Scientific Research C, JP23590549; Challenging Exploratory Research, JP26670064, JP16K15151; Young Scientists A, JP15H05644; Young Scientists B, JP18K14905).
Conflict of Interest
The authors declare no conflict of interest.
Supplementary Materials
The online version of this article contains supplementary materials.
REFERENCES
- 1) Henrickson KJ, Chanock R. Parainfluenza viruses. Clin. Microbiol. Rev., 16, 242–264 (2003).
- 2) Newman JT, Riggs JM, Surman SR, McAuliffe JM, Mulaikal TA, Collins PL, Murphy BR, Skiadopoulos MH. Generation of recombinant human parainfluenza virus type 1 vaccine candidates by importation of temperature-sensitive and attenuating mutations from heterologous paramyxoviruses. J. Virol., 78, 2017–2028 (2004).
- 3) Fry AM, Curns AT, Harbour K, Hutwagner L, Holman RC, Anderson LJ. Seasonal trends of human parainfluenza viral infections: United States, 1990–2004. Clin. Infect. Dis., 43, 1016–1022 (2006).
- 4) Counihan ME, Shay DK, Holman RC, Lowther SA, Anderson LJ. Human parainfluenza virus-associated hospitalizations among children less than five years of age in the United States. Pediatr. Infect. Dis. J., 20, 646–653 (2001).
- 5) Lucas JR, Haas EA, Masoumi H, Krous HF. Sudden death in a toddler with laryngotracheitis caused by human parainfluenza virus-1. Pediatr. Dev. Pathol., 12, 165–168 (2009).
- 6) Suzuki T, Portner A, Scroggs RA, Uchikawa M, Koyama N, Matsuo K, Suzuki Y, Takimoto T. Receptor specificities of human respiroviruses. J. Virol., 75, 4604–4613 (2001).
- 7) Fukushima K, Takahashi T, Ito S, Takaguchi M, Takano M, Kurebayashi Y, Oishi K, Minami A, Kato T, Park EY, Nishimura H, Takimoto T, Suzuki T. Terminal sialic acid linkages determine different cell infectivities of human parainfluenza virus type 1 and type 3. Virology, 464–465, 424–431 (2014).
- 8) Takaguchi M, Takahashi T, Hosokawa C, Ueyama H, Fukushima K, Hayakawa T, Itoh K, Ikeda K, Suzuki T. A single amino acid mutation at position 170 of human parainfluenza virus type 1 fusion glycoprotein induces obvious syncytium formation and caspase-3-dependent cell death. J. Biochem., 149, 191–202 (2011).
- 9) Merz DC, Scheid A, Choppin PW. Importance of antibodies to the fusion glycoprotein of paramyxoviruses in the prevention of spread of infection. J. Exp. Med., 151, 275–288 (1980).
- 10) Luque LE, Bridges OA, Mason JN, Boyd KL, Portner A, Russell CJ. Residues in the heptad repeat a region of the fusion protein modulate the virulence of Sendai virus in mice. J. Virol., 84, 810–821 (2010).
- 11) Pastey MK, Gower TL, Spearman PW, Crowe JE Jr, Graham BS. A RhoA-derived peptide inhibits syncytium formation induced by respiratory syncytial virus and parainfluenza virus type 3. Nat. Med., 6, 35–40 (2000).
- 12) Luque LE, Russell CJ. Spring-loaded heptad repeat residues regulate the expression and activation of paramyxovirus fusion protein. J. Virol., 81, 3130–3141 (2007).
- 13) Collins PL, Chanock RM, McIntosh K. Fields Virology. 3rd ed., Vol. 1 (Fields BN, Knipe DM, Howley PM, Chanock RM, MelnickJL, Monath TP, Roizman B, Straus SE ed.) Lippincott-Raven Publishers, Philadelphia, pp. 1205–1243 (1996).
- 14) Fukushima K, Takahashi T, Takaguchi M, Ueyama H, Ito S, Kurebayashi Y, Kawanishi T, McKimm-Breschkin JL, Takimoto T, Minami A, Suzuki T. Plaque formation assay for human parainfluenza virus type 1. Biol. Pharm. Bull., 34, 996–1000 (2011).
- 15) Suzuki T, Ikeda K, Koyama N, Hosokawa C, Kogure T, Takahashi T, Jwa Hidari KI, Miyamoto D, Tanaka K, Suzuki Y. Inhibition of human parainfluenza virus type 1 sialidase by analogs of 2-deoxy-2,3-didehydro-N-acetylneuraminic acid. Glycoconj. J., 18, 331–337 (2001).
- 16) Suzuki T, Sometani A, Yamazaki Y, Horiike G, Mizutani Y, Masuda H, Yamada M, Tahara H, Xu G, Miyamoto D, Oku N, Okada S, Kiso M, Hasegawa A, Ito T, Kawaoka Y, Suzuki Y. Sulphatide binds to human and animal influenza A viruses, and inhibits the viral infection. Biochem. J., 318, 389–393 (1996).
- 17) Huberman K, Peluso RW, Moscona A. Hemagglutinin-neuraminidase of human parainfluenza 3: role of the neuraminidase in the viral life cycle. Virology, 214, 294–300 (1995).
- 18) Yin H-S, Wen X, Paterson RG, Lamb RA, Jardetzky TS. Structure of the parainfluenza virus 5 F protein in its metastable, prefusion conformation. Nature, 439, 38–44 (2006).
- 19) Smith EC, Gregory SM, Tamm LK, Creamer TP, Dutch RE. Role of sequence and structure of the hendra fusion protein fusion peptide in membrane fusion. J. Biol. Chem., 287, 30035–30048 (2012).