MATERIALS AND METHODS
GeneralAll synthetic operations were conducted with a standard Schlenk technique under argon atmosphere. Flash column LC was performed using Kanto Chemical silica gel 60NO (spherical, 40–50 µm; Kanto Chemical Co., Inc., Tokyo, Japan). Analytical TLC was performed on Merck Kieselge1 60 F254 (0.25 mm) plates (Merck KGaA, Darmstadt, Germany). Visualization was accomplished with a 3% H2SO4 solution in methanol followed by heating.
ChemicalsUnless otherwise stated, commercially available chemicals were used as received. Pyridine (dehydrated), dichloromethane (super dehydrated), acetonitrile (super dehydrated), and N-hydroxysuccinimide (NHS) were purchased from FUJIFILM Wako Pure Chemical Corporation (Osaka, Japan). D-(+)-Mannose, 5-hexen-1-ol, tin (IV) chloride (approx. 1 mol/L in dichloromethane), and 1-(3-dimethylaminopropyl)-3-ethylcarbodiimide hydrochloride (EDC·HCl) were purchased from Tokyo Chemical Industry Co., Ltd. (Tokyo, Japan). Acetic anhydride, sodium periodate, and Celite® 535RVZ were purchased from Nacalai Tesque, Inc. (Kyoto, Japan). All other chemicals were of the highest grade available and were used without further purification.
OligonucleotidesPhosphodiester ODNs were purchased from Integrated DNA Technologies, Inc. (Coralville, IA, U.S.A.) and an Alexa Fluor 488-labeled ODN (Hexa-1) was purchased from Japan Bio Services Co., LTD. (Saitama, Japan). The oligonucleotide sequences were designed according to a previous work29) (Table 1). The ODN1668 with the same sequence as phosphorothioate CpG ODN 1668 and 5′-NH2-ODN1668 was used for Man-ODN1668 synthesis. The complementary sequence to the CpG motif (GAC GTT) in hexa-1 to -6 was designed to have a single base mismatch (AAC TTC) and no CG sequence to avoid TLR9 stimulation by these ODNs.
Table 1. Sequences of the ODNs
| Name | Sequence (5′ to 3′) | Base number |
|---|
| Hexa-1 | TAGCA GCACA TCAGG TTCTG AGCCT TGCTG CAAGC ATCAG GAACT TCATG GA | 52 |
| Hexa-2 | TGCAG CAAGG CTCAG ATCTG CTCAA GCCTG CAAGC ATCAG GAACT TCATG GA | 52 |
| Hexa-3 | TGCAG GCTTG AGCAG ACAGA GCCTT GAGCC TAAGC ATCAG GAACT TCATG GA | 52 |
| Hexa-4 | TAGGC TCAAG GCTCT GGAGG CTCTT AAGCT GCAGC ATCAG GAACT TCATG GA | 52 |
| Hexa-5 | GCAGC TTAAG AGCCT CAGAG CTTGG CATAG CAAGC ATCAG GAACT TCATG GA | 52 |
| Hexa-6 | TGCTA TGCCA AGCTC TACCT GATGT GCTGC TAAGC ATCAG GAACT TCATG GA | 52 |
| ODN1668 | TCCAT GACGT TCCTG ATGCT | 20 |
| 5′-NH2-ODN1668 | /5AmMC6/TCCAT GACGT TCCTG ATGCT | 20 |
All ODNs have a phosphodiester backbone. /5AmMC6/, 5′ amino modifier C6.
Proton and carbon NMR spectra (1H-NMR and 13C-NMR) were recorded on a JEOL JNM-ECZ400 (1H at 399.78 MHz and 13C at 100.53 MHz) spectrometer (JEOL Ltd., Tokyo, Japan), with solvent resonance as the internal standard (1H-NMR, CHCl3 at 7.26 ppm; 13C-NMR, CDCl3 at 77.0 ppm). The following 1H-NMR data were reported: integration, chemical shift multiplicity (s = singlet, d = doublet, t = triplet, q = quartet, m = multiplet), and coupling constants (Hz).
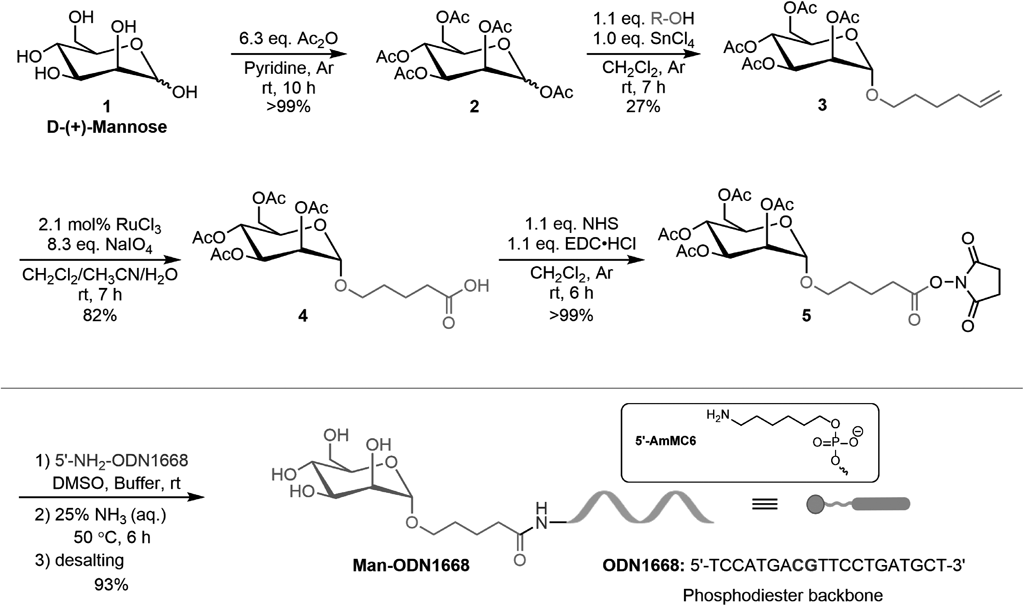
Synthesis of 5-Hexenyl 2,3,4,6-Tetra-O-acetyl α-D-Mannopyranoside (3)D-Mannose pentaacetate (2) was synthesized according to a previous report.30) Briefly, acetic anhydride (63.4 mmol, 6.47 mg, 6.0 mL, 6.34 equivalent (equiv.)) was added dropwise to a solution of D-(+)-mannose (10 mmol, 1.80 g) in pyridine (20 mL) at room temperature (r.t.). The reaction mixture was further stirred at r.t. for 10 h. The resulting mixture was quenched by methanol and concentrated with toluene in vacuo to yield a light-yellow oil (>99%). The crude residue was directly used for the glycosylation of 5-hexen-1-ol. After SnCl4 (1.0 mol/L in CH2Cl2, 0.5 mL, 1.0 equiv.) was added dropwise to a solution of compound 2 (0.53 mmol, 209.9 mg) in CH2Cl2 (7 mL), 5-hexen-1-ol (0.58 mmol, 58.4 mg, 70.0 µL, 1.1 equiv.) was immediately added to the reaction mixture with a vigorous stir. The reaction mixture was further stirred at r.t. for 5 h. The resulting mixture was quenched by sat. NaHCO3 (aq.), poured into a separatory funnel, and extracted three times with CH2Cl2. The organic layer was filtered through Celite® and concentrated. The crude residue was further purified on silica gel column using hexane and ethyl acetate (3 : 1) as eluent to yield the desired product in a colorless oil liquid (62.5 mg, 27%). 1H-NMR (400 MHz, CDCl3): δ = 5.79–5.69 (1H, m), 5.28 (1H, dd, J = 10.0, 3.4 Hz), 5.22 (1H, d, J = 9.9 Hz), 5.18–5.16 (1H, m), 4.98–4.88 (2H, m), 4.74 (1H, d, J = 1.7 Hz), 4.21 (1H, dd, J = 12.2, 5.3 Hz), 4.04 (1H, dd, J = 12.2, 2.4 Hz), 3.99–3.84 (1H, m), 3.63 (1H, dt, J = 12.1, 4.8 Hz), 3.40 (1H, dt, J = 11.8, 4.8 Hz), 2.09 (3H, s), 2.05–1.99 (8H, m), 1.93 (3H, s), 1.60–1.53 (2H, m), 1.44–1.38 (2H, m). 13C-NMR (101 MHz, CDCl3): δ = 170.42, 169.87, 169.69, 169.56, 138.20, 114.67, 97.38, 69.52, 68.96, 68.26, 68.11, 66.07, 62.35, 33.19, 28.49, 25.16, 20.71, 20.54, 20.51, 20.50 ppm. MS (electrospray ionization (ESI)+): Calcd for C20H30O10Na+ [M + Na]+: 453.1731. Found: m/z 453.1733.
Synthesis of 5-[(Tetra-O-acetyl α-D-Mannopyranosyl)oxyl] Pentanoic Acid (4)RuCl3·H2O (aq.) (1.29 mg/mL, 0.48 mL; 3 µmol, 0.62 mg, 2.1 mol %) was added to a solution of compound 3 (0.145 mmol, 62.5 mg) in CH2Cl2/CH3CN (1 : 1, 0.64 mL). Thereafter, NaIO4 (0.6 mmol, 127.9 mg, 4.15 equiv.) was immediately added to the reaction mixture with a vigorous stir. After 2 h, more NaIO4 (0.6 mmol, 127.9 mg, 4.15 equiv.) was added to the reaction mixture, which was further stirred at r.t. for 2 h. The resulting mixture was diluted with H2O, poured into a separatory funnel, and extracted three times with CH2Cl2. The organic layer was dried over MgSO4 and concentrated in vacuo to yield a dark oil liquid. The crude residue was directly used for the subsequent reaction (53.5 mg, 82%, crude yield). 1H-NMR (400 MHz, CDCl3): δ = 5.33 (1H, dd, J = 10.0, 3.4 Hz), 5.28 (1H, d, J = 9.8 Hz), 5.24–5.22 (1H, m), 4.80 (1H, d, J = 1.5 Hz), 4.27 (1H, dd, J = 12.2, 5.4 Hz), 4.10 (1H, dd, J = 12.2, 2.4 Hz), 3.99–3.95 (1H, m), 3.71 (1H, dt, J = 11.1, 5.0 Hz), 3.47 (1H, dt, J = 11.0, 5.0 Hz), 2.42–2.39 (2H, m), 2.15 (3H, s,), 2.10 (3H, s), 2.04 (3H, s), 1.99 (3H, s), 1.75–1.67 (4H, m). 13C-NMR (101 MHz, CDCl3): δ = 178.33, 170.70, 170.12, 169.98, 169.77, 97.57, 69.62, 69.11, 68.47, 67.89, 66.17, 62.52, 33.37, 28.53, 21.31, 20.89, 20.72, 20.69 ppm. MS (ESI+): Calcd for C19H28O12Na+ [M + Na]+: 471.1473. Found: m/z 471.1473.
Synthesis of 2,5-Dioxopyrrolidin-1-yl 5-[(Tetra-O-acetyl α-D-Mannopyranosyl)oxyl] Pentanate (5)1-(3-Dimethylaminopropyl)-3-ethylcarbodiimide hydrochloride (0.125 mmol, 24.0 mg, 1.05 equiv.) was added to a solution of compound 4 (0.119 mmol, 53.5 mg) and N-hydroxysuccinimide (0.125 mmol, 14.4 mg, 1.05 equiv.) in CH2Cl2 (1 mL), which was stirred at r.t. for 6 h. The resulting mixture was quenched by H2O, poured into a separatory funnel, and extracted three times using CH2Cl2. The organic layer was washed with brine, dried over Na2SO4, and concentrated in vacuo. The crude residue was directly used for modification of CpG1668 (65.4 mg, >99%, crude yield, dark oil liquid). 1H-NMR (400 MHz, CDCl3): δ = 5.29 (1H, dd, J = 10.0, 3.4 Hz), 5.24 (1H, d, J = 9.8 Hz), 5.21–5.19 (1H, m), 4.78 (1H, d, J = 1.5 Hz), 4.24 (1H, dd, J = 12.2, 5.3 Hz), 4.06 (1H, dd, J = 12.2, 2.2 Hz), 3.92–3.96 (1H, m), 3.71 (1H, dt, J = 11.3, 4.9 Hz), 3.47 (1H, dt, J = 11.2, 4.9 Hz), 2.80 (4H, s), 2.64 (2H, t, J = 7.0 Hz), 2.12 (3H, s), 2.06 (3H, s), 2.01 (3H, s), 1.95 (3H, s), 1.86–1.78 (2H, m), 1.75–1.69 (2H, m) ppm. 13C-NMR (101 MHz, CDCl3): δ = 170.56, 169.95, 169.75, 169.66, 169.08, 168.23, 97.48, 69.49, 68.97, 68.40, 67.47, 66.10, 62.42, 30.47, 28.11, 25.50, 21.33, 20.80, 20.65, 20.61, 20.59 ppm. MS (ESI+): Calcd for C23H31NO14Na+ [M + Na]+: 568.1637. Found: m/z 568.1645.
Synthesis of Mannosylated ODN1668 (Man-ODN1668)After 5′-NH2-ODN1668 (20 nmol, 1 mM in distilled H2O, 20 µL) was added to a mixture of Na2CO3–NaHCO3 buffer (0.33 M, pH = 9, 60 µL) and dimethyl sulfoxide (DMSO) (64 µL), compound 5 (20 µL, 0.5 M in DMSO, 40 µL) was added to the mixture. After moderate mixing of all the reagents using a vortex mixer and centrifugation, the mixture was stored overnight at r.t. for a further reaction. The reactant was purified with a Zeba spin desalting column (7K MWCO, Thermo Fisher Scientific Inc., U.S.A.) according to the manufacturer’s protocol. Thereafter, it was lyophilized to yield a white powder which was resolved in distilled H2O. The formation of Man-ODN1668 was confirmed by matrix assisted laser desorption ionization-time of flight MS (MALDI-TOF MS, JMS-S3000, JEOL Ltd.). MS Calcd: 6501.4. Found: 6501.1. Furthermore, its yield of 85% was confirmed by Nanodrop 2000/2000c (Thermo Fisher Scientific Inc., U.S.A.).
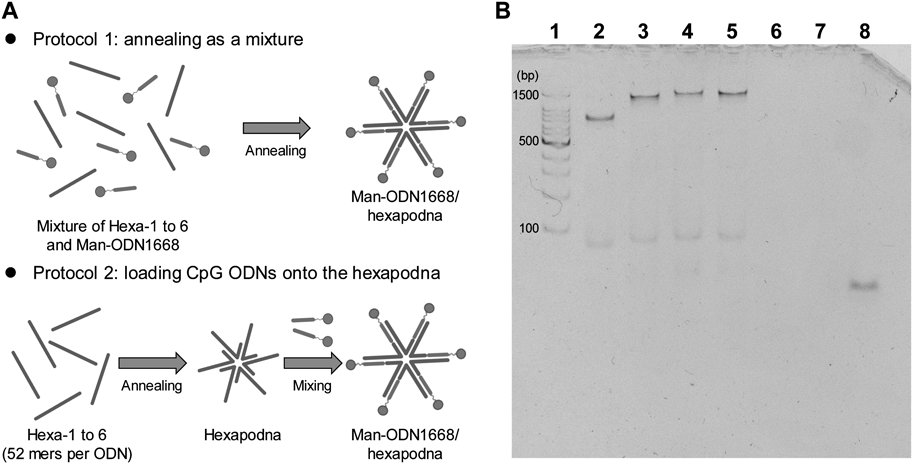
Preparation of PolypodnaAll ODNs involved in polypodna formation were mixed in appropriate molar ratios, heated to 95°C, and then gently cooled to 4°C. The preparation protocol was reported previously.14)
Polyacrylamide Gel Electrophoresis (PAGE)The formation of hexapodnas was confirmed by PAGE (6% polyacrylamide gel), which was carried out at 200 V for 30 min at rt. A total of 50 ng of each DNA sample was added to the gel for PAGE. The 100-bp DNA ladder was purchased from TaKaRa (Tokyo, Japan). ODNs were visualized by staining with ethidium bromide (EtBr; Nippon Gene Co., Ltd., Tokyo, Japan) and observed with the LAS4000 imaging system (FUJIFILM, Tokyo, Japan).
Measurement of Melting TemperatureHexapodnas were diluted in 150 mM NaCl to 250 µg/mL. Melting temperature (Tm) was measured with a spectrophotometer (JASCO J-730, JASCO, Tokyo, Japan) in a quartz cell with an optical path length of 1 mm (JASCO). All samples were scanned from 20°C to 95°C at a rate of 1°C/min and the absorbances were recorded per 0.5°C.
AnimalsC57BL/6N mice (female, 6-week-old) were purchased from Sankyo Labo Service Co., Inc. (Tokyo, Japan). All animal experiments were approved by the Animal Research Committee of the Faculty of Pharmaceutical Sciences, Tokyo University of Science.
Isolation and Culture of Mouse Peritoneal MacrophagesThe elicited macrophages were harvested from the peritoneal cavity of C57BL/6N mice 3 d after an intraperitoneal injection of 2 mL of 2.9% thioglycolate medium (Nissui Pharmaceutical Co., Ltd., Tokyo, Japan). After cells were washed, they were suspended in RPMI 1640 (Nissui Pharmaceutical Co., Ltd.) supplemented with 10% heat-inactivated fetal bovine serum (Gibco, Thermo Fisher Scientific Inc., Waltham, MA, U.S.A.), penicillin G (100 U/mL), streptomycin (100 µg/mL) and L-glutamine (2 mmol/L) (FUJIFILM Wako Pure Chemical Corporation) in a humidified incubator containing 5% CO2 at 37°C. Thereafter, the cells were plated on a 10-cm dish. After incubation for 2 h, nonadherent macrophages were removed via washing with culture medium. As the attached cells were regarded as peritoneal macrophages, they were harvested for further experiments.
Culture of Mouse Macrophage-Like RAW264.7 Cells and J774.1 CellsMouse macrophage-like RAW264.7 cells and J774.1 cells were cultured in RPMI 1640 (Nissui Pharmaceutical Co., Ltd.) supplemented with 10% heat-inactivated fetal bovine serum (Gibco, Thermo Fisher Scientific Inc.), penicillin G (100 U/mL), streptomycin (100 µg/mL) and L-glutamine (2 mmol/L) (FUJIFILM Wako Pure Chemical Corporation) in a humidified incubator containing 5% CO2 at 37°C.
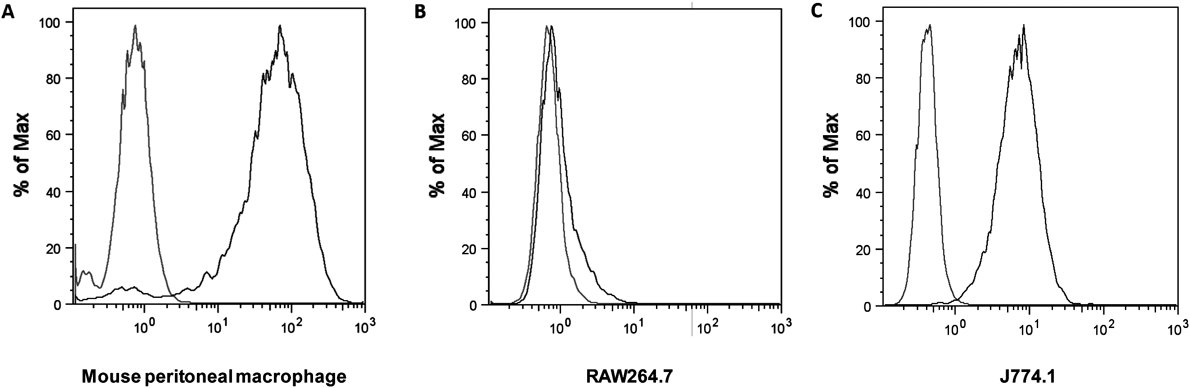
Identification of the Expression of MRThe expression of MR on mouse peritoneal macrophages, RAW264.7 cells and J774.1 cells was identified by direct immunofluorescence staining using anti-CD206 antibody (rat anti mouse CD206: Alexa Fluor 488, Bio-Rad Laboratories Inc., Hercules, CA, U.S.A.) following the manufacturer’s protocol. Briefly, 1 × 106 cells suspended in 90 µL cold (4°C) phosphate buffered saline (PBS)/bovine serum albumin (BSA) buffer were added with 10 µL anti-CD206 antibody, and the mixture was incubated for 1 h at 4°C in dark. Then, the cells were centrifuged, washed with PBS/BSA, and resuspended in 200 µL cold PBS. The fluorescence intensity of the cells was determined using a flow cytometer (Gallios Flow Cytometer; Beckman Coulter, Miami, FL, U.S.A.). Data were analyzed using FlowJo software (version 8.8.4; Beckman Coulter).
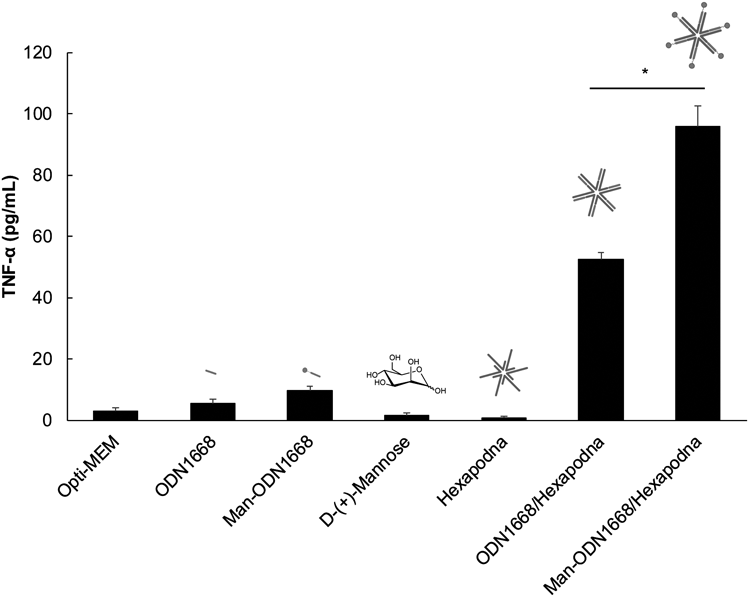
Tumor Necrosis Factor (TNF)-α Release from Peritoneal Macrophages and RAW264.7 CellsCells were seeded into a 96-well plate at a density of 1 × 105 cells/well (mouse peritoneal macrophages) or 5 × 104 cells/well (RAW264.7 cells) and incubated for 24 h before treatment. After aspiration of the supernatant, the DNA samples diluted with Opti-modified Eagle’s medium (Opti-MEM, Gibco, Thermo Fisher Scientific) were added to cells at an ODN1668 concentration of 1.2 µM (mouse peritoneal macrophages) or 0.6 µM (RAW264.7 cells). Thereafter, the cells were further incubated for 8 h at 5% CO2 and 37°C. The supernatant was then collected and used to determine the concentrations of TNF-α by enzyme-linked immunosorbent assay (ELISA), following the manufacturer’s protocol (BioLegend, Inc., San Diego, CA, U.S.A.).
IL-6 Release from J774.1 CellsJ774.1 cells (1 × 104 cells/well) were seeded into a 96-well plate and incubated at 37°C for 24 h before treatment. DNA samples diluted with Opti-MEM were added to cells at an ODN1668 concentration of 4.2 nM. Thereafter, the cells were further incubated for 8 h at 5% CO2 and 37°C. The supernatant was then collected and used to determine the concentrations of interleukin (IL)-6 by ELISA following the manufacturer’s protocol (Thermo Fisher Scientific Inc.).
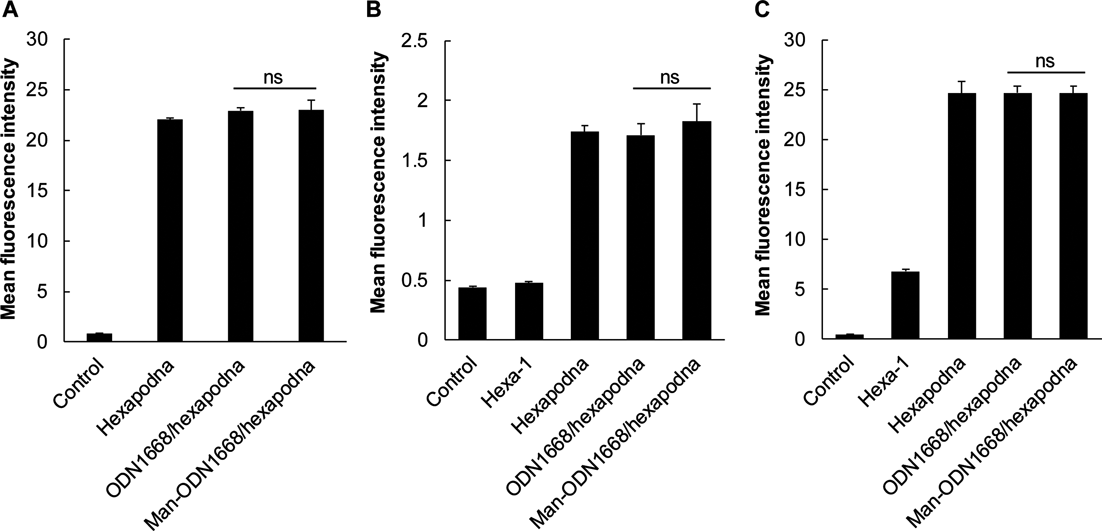
Cellular UptakeAlexa Fluor 488-labaled Hexa-1 was used for the construction of Alexa Fluor 488-labaled DNA samples. Cells were seeded into 24-well plates at a density of 1 × 105 cells/well and incubated for 24 h before treatment. After aspiration of the supernatant, the DNA samples diluted with Opti-MEM were added to cells at a DNA concentration of 1.2 µM (mouse peritoneal macrophages), 0.6 µM (RAW264.7 cells) or 4.2 nM (J774.1 cells). Thereafter, the cells were further incubated for 2 h at 5% CO2 and 37°C. Cells were then washed twice with PBS and harvested. Then, fluorescence intensity of the cells was determined using a flow cytometer (Gallios Flow Cytometer). Data were analyzed using FlowJo software (version 8.8.4).