Abstract
Proton pumping ATPases, both F-type and V/A-type ATPases, generate ATP using electrochemical energy or pump protons/sodium ions by hydrolyzing ATP. The enzymatic reaction and proton transport are coupled through subunit rotation, and this unique rotational mechanism (rotational catalysis) has been intensively studied. Single-molecule and thermodynamic analyses have revealed the detailed rotational mechanism, including the catalytically inhibited state and the roles of subunit interactions. In mammals, F- and V-ATPases are involved in ATP synthesis and organelle acidification, respectively. Most bacteria, including anaerobes, have F- and/or A-ATPases in the inner membrane. However, these ATPases are not believed to be essential in anaerobic bacteria since anaerobes generate sufficient ATP without oxidative phosphorylation. Recent studies suggest that F- and A-ATPases perform indispensable functions beyond ATP synthesis in oral pathogenic anaerobes; F-ATPase is involved in acid tolerance in Streptococcus mutans, and A-ATPase mediates nutrient import in Porphyromonas gingivalis. Consistently, inhibitors of oral bacterial F- and A-ATPases, such as phytopolyphenols and bedaquiline, strongly diminish growth and survival. Herein, we discuss rotational catalysis of bacterial F- and A-ATPases, and discuss their physiological roles, focusing on oral bacteria. We also review the effects of ATPase inhibitors on the growth and survival of oral pathogenic bacteria. The features of the catalytic mechanism and unique physiological roles in oral bacteria highlight the potential for proton pumping ATPases to serve as targets for oral antimicrobial agents.
1. INTRODUCTION
The oral ecosystem comprises the oral flora and the oral microbiome.1) Some oral pathogens are responsible for dental caries and periodontal disease, resulting in loss of teeth.2) In addition, chronic oral infection with these microorganisms is associated with systemic diseases such as cancers, type 2 diabetes, and Alzheimer’s disease.3–5) However, antimicrobial agents have rarely been used for therapy or prevention of dental caries and periodontal disease, since continuous use of these drugs induces drug-resistant strains and disrupts the oral microbiome. Therefore, there is a need to identify novel targets in pathogens to develop oral antimicrobial agents.
Proton pumping ATPases are membrane-embedded, multisubunit enzymes coupling proton or sodium ion transport and ATP synthesis or hydrolysis through subunit rotation.6) They are classified into F-type and V/A-type ATPases.6) F-type ATPase (F-ATPase, also known as ATP synthase) is found in chloroplasts, mitochondria, and bacterial membranes.7) Vacuolar-type ATPase (V-ATPase) is present in eukaryote cells, localized in endomembrane organelles such as vacuoles, lysosomes, endosomes, and Golgi apparatus, as well as the plasma membrane.8) Archaeal type ATPase (A-ATPase) in bacterial membrane appears to be the ancestors of V-ATPase.9) Therefore, A-ATPase is also called V/A-ATPase.
The unique structures and rotational mechanism of proton pumping ATPases have been intensively studied.10–13) Specifically, single-molecule studies on rotational catalysis by proton pumping ATPases have revealed their dynamics, torque generation, and intrinsic features such as stochastic rotation rate and catalytically inhibited state, none of which could be determined by bulk phase analysis.14–17) Studies involving single-molecule assays, crystal structures, and inhibitors of proton pumping ATPases have revealed the importance of subunit interactions for smooth rotation.18–21)
Eukaryotic F- and V-ATPases are involved in ATP synthesis and organelle acidification, respectively.6) F- and V-ATPases are essential in mammals since their deletion is embryonic lethal.22,23) Most bacteria, including anaerobes, possess F-ATPase and/or A-ATPase in the inner membrane. However, these ATPases are believed to be unessential for energy metabolism in anaerobic bacteria because these organisms generate sufficient ATP through anaerobic glycolysis without depending on ATP synthesis via oxidative phosphorylation.24,25) Recently, it was reported that F- and A-ATPases in oral pathogenic anaerobes play indispensable roles in acid tolerance by secreting protons from the cytoplasm, and in amino acid import by generating a proton gradient.26–28) These findings suggest that F- and A-ATPases could serve as potential targets for antimicrobial agents against oral pathogens.
Consistently, proton pumping ATPase inhibitors diminish the growth and survival of oral pathogenic bacteria including Streptococcus mutans and Porphyromonas gingivalis.27,28) Most inhibitors are toxic to both bacteria and mammals, probably due to their structural similarity, whereas phytopolyphenols and the antituberculosis drug bedaquiline are less toxic to mammals.20,29–32) This is possibly because bedaquiline selectively affects bacterial F-ATPase, and phytopolyphenols are rapidly metabolized and/or less readily absorbed in mammals.33,34) Therefore, we predict that these less toxic inhibitors could serve as seeds for novel antidental caries and antiperiodontitis agents.
Herein, we focus on bacterial F- and A-type proton pumping ATPases. We review the mechanism underlying rotational catalysis of F- and A-ATPases, discuss the physiological roles of these ATPases in oral pathogenic bacteria, and describe the effects of ATPase inhibitors on the growth and survival of pathogenic bacteria. Understanding the chemomechanical coupling mechanism of proton pumping ATPases will provide pharmacophores for these ATPases, and accelerate the discovery of seed compounds for oral pathogen-specific antimicrobial agents.
2. ROTATIONAL CATALYSIS OF PROTON PUMPING ATPASES
2.1. Rotational Catalysis of F-Type ATPaseF-ATPase (Fig. 1A) consists of a catalytic F1 sector (α3β3γδε) and a transmembrane proton pumping FO sector (ab2c8–14).35–38) The rotor complex composed of γεc8–14 subunits rotates against the stator complex built from α3β3δab2 subunits, which couples catalysis and proton translocation.7) Rotational catalysis of F-ATPase has been studied by single-molecule observation and crystal structure analysis.7) The γ subunit in Escherichia coli F1 rotates relative to the α3β3 hexamer in distinct 120° steps during ATP hydrolysis39) (Fig. 1B). During 360° rotation of the γ subunit, the β subunit sequentially passes through three different conformations: βE (no nucleotide), βTP (bound ATP), and βDP (bound ADP; Figs. 1B, C).40,41) In each 120° step, ATP binds to βE, βTP hydrolyzes ATP to ADP + Pi (inorganic phosphate), and Pi and ADP are released from βDP (Fig. 1C; β subunit highlighted by an orange line).40,41)
To analyze the rotational mechanism of F1, 40 or 60 nm gold beads were attached to the γ subunit of the F1 sector immobilized on a glass surface through a histidine (His)-tag attached to the α subunit.19–21,39,41–44) Rotation of the attached gold beads relative to the α3β3 sector was observed under laser light illumination.43) The viscous drag of the probe was low enough to observe high-speed rotation of the γ subunit (approx. 500 rotations per second under Vmax conditions).43) The rotation rate was stochastic, and, interestingly, the γ subunit underwent continuous rotation (active state) and pausing (catalytically inhibited state) every approx. 1 s.43,44) The ε subunit, an intrinsic inhibitor of F-ATPase, lowers the rotation rate in the active state and extends the duration of the inhibited state.19,44)
We performed thermodynamic analysis of single-molecule rotation by observing temperature-dependent rotation of the γ subunit.20,41,44) We found that the activation energy of catalytic steps was similar, suggesting there is no obvious rate-limiting step throughout the rotation.41) In other words, γ subunit rotation mediates the highly cooperative reaction mechanism and lowers energy barriers for ATP hydrolysis/synthesis by F-ATPase.41)
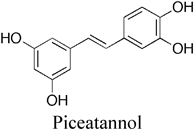
2.2. Subunit Interactions of F-ATPaseSingle-molecule assays together with thermodynamic and crystal structure analyses revealed the importance of subunit interactions.20,41,44) N- and C-terminal regions of the γ subunit interact with βDP and βTP, respectively45) (Fig. 1D). The γM23K mutation (E. coli F1 numbering) results in an additional hydrogen bond between βDP and the N-terminal region of the γ subunit.46) Piceatannol, an F-ATPase inhibitor, binds to the interface between βTP and the C-terminal region of the γ subunit.47) Thermodynamic analysis indicated that the γM23K mutation and piceatannol synergistically increased the activation energy in the rate-limiting step by preventing the β subunit from triggering γ rotation, suggesting that interactions between β and γ subunits at both interfaces influence the rate-limiting transition state.20,41)
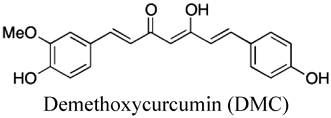
The region including βArg398 of βDP interacts with that including αGlu402 of αDP45) (Fig. 1D). Mutational and molecular docking studies suggest that the F-ATPase inhibitors curcumin and citreoviridin bind to the region around βArg398 and disrupt the interaction between α and β subunits.21) These inhibitors prevent conformational transmission of the β subunit,21) suggesting that the α–β interaction plays a role in the β subunit conformational changes required for triggering γ subunit rotation. These studies revealed that interactions at the α–β and β–γ interfaces of F-ATPase are crucial for smooth progression through the reaction cycle, and suggest that compounds binding to these interfaces may inhibit rotational catalysis.20,41) Therefore, these interfaces could serve as pharmacophores for in silico screening of F-ATPase inhibitors.7,21)
2.3. Rotational Catalysis of A-ATPaseA-ATPase consists of a catalytic A1 (A3B3DF) sector, a transmembrane proton pumping AO sector (acxd), and a stalk sector (E2G2), but the number of c subunits varies between bacterial species48,49) (Fig. 2A). The rotor complex of the DFcxd subunits rotates against the stator complex of the A3B3E2G2a subunits.48,49) The structure, subunit composition, and rotational catalysis of the enzyme resemble those of F-ATPase.12,50) The D subunit in A1 rotates relative to the A3B3 hexamer in distinct 120° steps51,52) (Fig. 2B). Similar to F-ATPase, A-ATPase is characterized by a stochastic rotation rate and a catalytically inhibited state,51,53) but A-ATPase of Enterococcus hirae does not adopt the inhibited state.51) Amino acid residues in the catalytic sites of A and B subunits are similar to those in β and α subunits of F-ATPase.54,55) During rotation, pairs of A and B subunits pass through three different conformations, empty, bound, and tightly bound, corresponding to αEβE, αTPβTP, and αDPβDP of F-ATPase, respectively50) (Figs. 1B, 2B). According to the crystal structure of the enzyme, the N- and C-terminal helices of the D subunit interact with the A subunit to trigger rotation, suggesting that subunit interactions mediate rotational catalysis.50) Further studies including thermodynamic analysis will likely reveal the detailed rotational mechanism of A-ATPase and the essential interfaces that may serve as pharmacophores for specific inhibitors.
3. PHYSIOLOGICAL ROLES OF PROTON PUMPING ATPASES IN BACTERIA
3.1. Physiological Roles of F-ATPaseF-ATPase in the bacterial inner membrane is essential for the production of ATP in aerobic bacteria such as Mycobacterium tuberculosis, the etiological agent of tuberculosis.56)M. tuberculosis lowers its metabolism when entering the nongrowing (dormant) state under hypoxic conditions, and becomes recalcitrant to most antituberculous drugs.56) Importantly, F-ATPase inhibitors decrease bacterial growth in the optimal replicating state, and diminish survival in the hypoxic dormant state.57) These studies suggest that F-ATPase plays an important role in maintaining low ATP levels in the dormant state in M. tuberculosis, hence inhibitors could decrease bacterial survival in the dormant state.
On the other hand, F-ATPase is believed to be unessential in anaerobic bacteria since they can bypass the requirement for oxidative phosphorylation when grown on carbohydrates. Consistently, knockout of F-ATPase genes does not significantly decrease the growth of anaerobic bacteria such as E. coli and Staphylococcus aureus under nutrient-rich conditions.24,25) However, recent studies indicate that F-ATPase is essential in pathogenic oral streptococci such as S. mutans and Streptococcus anginosus, even though they lack a respiratory chain for oxidative phosphorylation.58)
S. mutans is considered an etiological agent of dental caries; this bacterium produces organic acids through metabolism of dietary carbohydrates, which cause demineralization of the tooth enamel surface.59) Highly evolved acid tolerance mechanisms support bacterial growth and survival in the acidic microenvironment generated by the bacterium.60) F-ATPase likely contributes to an acid tolerance mechanism since expression of F-ATPase and membrane ATPase activity of S. mutans are elevated under acidic conditions.61) Additionally, we showed that F-ATPase inhibitors significantly decreased the growth rate and colony-forming ability of S. mutans under acidic conditions, but they had no effect under neutral conditions.27) These findings suggest that F-ATPase is involved in acid tolerance of S. mutans, probably by secreting protons from the cytoplasm.
S. anginosus is a pathogenic streptococcus found in the human oral cavity, and gastrointestinal and genital tracts.62) This bacterium causes serious purulent abscesses in various tissues, and subacute infective endocarditis,63,64) and it has been linked to gastrointestinal and esophageal cancers.65–67)S. anginosus has high acid tolerance and membrane ATPase activity, similar to S. mutans.26) Recent studies and our unpublished data showed that knockout of the F-ATPase β subunit and F-ATPase inhibitors significantly decreased acid tolerance of S. anginosus, suggesting that the enzyme also plays a role in acid tolerance.26)
By contrast, low pathogenic oral streptococci, such as S. sanguis, have lower acid tolerance and lower membrane ATPase activity than S. mutans and S. anginosus.26,68) This correlation between expression of F-ATPase, acid tolerance, and pathogenicity in oral streptococci implies that F-ATPase is involved in virulence by secreting protons from cytoplasm. Therefore, the enzyme could be a target for prevention of dental caries and diseases related to oral pathogenic streptococci.
3.2. Physiological Roles of A-ATPaseA-ATPase is found in the membrane of archaea such as Methanosarcina mazei Gö1 and Thermus thermophilus, where it functions in ATP synthesis.48) Some nonarchaeal anaerobes also possess this enzyme. Recent studies suggest that A-ATPase is essential for functions other than ATP synthesis in nonarchaeal oral pathogenic bacteria including P. gingivalis and Enterococcus faecalis.
P. gingivalis is a major causative agent of periodontitis.69) This bacterium is asaccharolytic, and utilizes amino acids and oligopeptides as energy sources.70) P. gingivalis lacks F-ATPase but expresses A-ATPase in the bacterial membrane.28) Proton pumping ATPase inhibitors significantly reduced ATPase activity in the bacterial membrane, where A-ATPase is located, and decreased bacterial growth.28) These findings suggest that the enzyme is essential for growth of P. gingivalis.28) Since the bacterium imports amino acids and oligopeptides through proton-dependent oligopeptide transporters (POTs), A-ATPase likely plays a role in generating the proton gradient for nutrient transport.71)
E. faecalis, found in a variety of environments including the root canal, causes infectious diseases such as endocarditis and endodontic failure.72,73) Interestingly, this bacterium possesses both F- and A-ATPases. E. faecalis F-ATPase synthesizes ATP, and A-ATPase functions in sodium extrusion from cytoplasm under high salt or alkaline conditions.74)
As described above, A-ATPase in nonarchaeal anaerobes mediates nutrient transport and salt tolerance, whereas it functions in central metabolism in archaea. It is likely that nonarchaeal bacteria adopted A-ATPase for various purposes during evolution. Since mammals do not have A-ATPase, it is a potential therapeutic target for oral infectious diseases such as periodontitis.
4. INHIBITORS OF BACTERIAL PROTON PUMPING ATPASES
There are various types of F-ATPase inhibitors, including dicyclohexylcarbodiimide (DCCD), azide, chloro-4-nitrobenzo-2-oxa-1,3-diazo1e (NBD-Cl), and aurovertin. Most inhibit both bacterial and mitochondrial F-ATPase since the structures of these enzymes are similar.31,75–77) Therefore, F-ATPase inhibitors are presumed to be toxic to mammals and unsuitable as antimicrobial agents. However, recent studies revealed that bedaquiline selectively affects bacterial F-ATPase, hence it is less toxic to mammals than previously expected.29)
On the other hand, only a few compounds including diethylstilbestrol and phytopolyphenols have been reported to inhibit A-ATPase.28,78) As described above, mammals do not possess this enzyme, and the structure of mammalian F- and V-ATPases is different from that of bacterial A-ATPase.28) Thus, A-ATPase inhibitors would presumably selectively kill bacteria.
4.1. BedaquilineBedaquiline, a diarylquinoline derivative, selectively binds to the membrane-spanning part of the c subunit of mycobacterial F-ATPase79) and functions as a H+/K+ ionophore80) (Table 1). It is likely that bedaquiline is accumulated in the mycobacterial membrane by binding to F-ATPase, which disrupts transmembrane proton and potassium gradients, resulting in inhibition of ATP synthesis.80) Amino acid residues essential for binding to bedaquiline are not conserved in mycobacterial and mitochondrial enzymes,29,79) explaining the selective binding of bedaquiline to mycobacterial F-ATPase. As described above, M. tuberculosis enters a nongrowing dormant state that is not very susceptible to known antituberculous drugs.56) Unlike these drugs, bedaquiline potently kills the bacterium in both the optimal replicating state and the dormant state57,81) (Table 1). Therefore, bedaquiline has recently been approved in 109 countries for treating multidrug-resistant tuberculosis.82)
Bedaquiline also decreases the growth and survival of S. mutans under acidic conditions, although the IC50 value for S. mutans is approx. 100 times higher than that for M. tuberculosis27,29) (Table 1). More recently, it was reported that bedaquiline analogs and other compounds binding to the bedaquiline binding site display more potent antituberculosis activity and/or less toxicity to mammals than bedaquiline.83,84) These inhibitors could be potential candidates for antidental caries agents.
4.2. PhytopolyphenolsPhytopolyphenols are a group of natural products found in plants including fruits and vegetables.85) Our group and others have shown that E. coli F-ATPase is inhibited by phytopolyphenols, such as piceatannol, resveratrol, quercetin, curcumin, thymoquinone, safranal, and olive phenolics20,32,86–88) (Table 1). As described above, the interface between the β subunit and the C-terminal regions of the γ subunit is important for triggering rotation of the γ subunit,23) and piceatannol, resveratrol, and quercetin bind to this interface.47) Mutagenesis and molecular docking studies suggest that curcumin and a 3′-demethoxy analog (demethoxycurcumin; DMC) interact with residues in interface between the α and β subunits of F1.21,89) Piceatannol, curcumin, and DMC decrease E. coli growth by inhibiting oxidative phosphorylation, consistent with their inhibitory effects on F1 ATPase activity20,30,32) (Table 1). These phytopolyphenols also inhibit S. mutans F-ATPase, and reduce bacterial growth and survival only under acidic conditions27) (Table 1), suggesting that they lower acid tolerance of pathogenic streptococci.
Curcumin lowers ATPase activity in the P. gingivalis membrane, where A-ATPase is located, and thereby inhibits bacterial growth28) (Table 1). Piceatannol and its analogs have similar chemical structures to diethylstilbestrol, an A-ATPase inhibitor, and they strongly decrease bacterial growth (Table 1). These findings suggest that phytopolyphenols exhibit antimicrobial activity against P. gingivalis by inhibiting A-ATPase.
Phytopolyphenols such as piceatannol and curcumin also inhibit ATPase activity in the mitochondrial membrane, where F-ATPase is located,90) and these compounds are not toxic to mammals, unlike other F-ATPase inhibitors. This is probably because piceatannol and its analogs are rapidly metabolized and eliminated in mammals.34) In addition, curcumin and its analogs are poorly soluble and unstable in water.33) It is unlikely that these phytopolyphenols inhibit F-ATPase in mammalian tissues due to their low bioavailability. On the other hand, these compounds are expected to inhibit F-ATPase in oral pathogenic bacteria since they can be used at high concentrations through oral administration without absorption, distribution, metabolism, and excretion (ADME) restrictions. Therefore, these phytopolyphenols could be potential seeds for novel antidental caries or periodontal agents.
5. CONCLUSION
Proton pumping ATPases in most organisms are involved in ATP synthesis, but these enzymes are presumed to be unessential in anaerobes that can grow without oxidative phosphorylation. Recent studies indicated that these enzymes in oral pathogenic anaerobic bacteria play indispensable roles in acid tolerance and nutrient transport, rather than ATP synthesis. Bedaquiline and phytopolyphenols affect proton pumping ATPases in these bacteria and inhibit the bacterial growth rate, and they are less toxic to mammals.27–29,32) Therefore, these compounds could be seeds for antimicrobial agents against oral pathogenic bacteria.
Studies on bacterial proton pumping ATPases based on single-molecule observations have revealed the essential mechanism of rotational catalysis, including the roles of the β−γ and α−β interfaces.7,20,21,41) Piceatannol and curcumin bind to these interfaces and inhibit proton pumping ATPases,20,21,47) suggesting that these interfaces may be applicable as pharmacophores for in silico screening of pathogen selective inhibitors.
Proton pumping ATPases are composed of many subunits, and various subunit interfaces essential for rotational catalysis have been identified.7) This implies that inhibitors binding to different interfaces of the enzymes could be developed. Antimicrobial resistance has become a worldwide problem. Cocktails of such inhibitors could more effectively kill bacteria than single administration and suppress the emergence of drug-resistant microbes.
Acknowledgments
Dr. M. Nakanishi-Matsui is thanked for helpful advice and critical discussion. This work was supported in part by Japan Society for the Promotion of Science (JSPS) KAKENHI Grants (JP18K06629, JP21K06546, and 21H02627), and by the Keiryokai Research Foundation (Grant No. 142).
Conflict of Interest
The author declares no conflict of interest.
REFERENCES
- 1) Samaranayake L, Matsubara VH. Normal oral flora and the oral ecosystem. Dent. Clin. North Am., 61, 199–215 (2017).
- 2) Valm AM. The Structure of dental plaque microbial communities in the transition from health to dental daries and periodontal disease. J. Mol. Biol., 431, 2957–2969 (2019).
- 3) Olsen I, Singhrao SK. Can oral infection be a risk factor for Alzheimer’s disease? J. Oral Microbiol., 7, 29143 (2015).
- 4) Li X, Kolltveit KM, Tronstad L, Olsen I. Systemic diseases caused by oral infection. Clin. Microbiol. Rev., 13, 547–558 (2000).
- 5) Han YW, Wang X. Mobile microbiome: oral bacteria in extra-oral infections and inflammation. J. Dent. Res., 92, 485–491 (2013).
- 6) Futai M, Nakanishi-Matsui M, Okamoto H, Sekiya M, Nakamoto RK. Rotational catalysis in proton pumping ATPases: from E. coli F-ATPase to mammalian V-ATPase. Biochim. Biophys. Acta BBABioenerg., 1817, 1711–1721 (2012).
- 7) Nakanishi-Matsui M, Sekiya M, Futai M. ATP synthase from Escherichia coli: mechanism of rotational catalysis, and inhibition with the ε subunit and phytopolyphenols. Biochim. Biophys. Acta BBABioenerg., 1857, 129–140 (2016).
- 8) Futai M, Sun-Wada GH, Wada Y, Matsumoto N, Nakanishi-Matsui M. Vacuolar-type ATPase: a proton pump to lysosomal trafficking. Proc. Jpn. Acad. Ser. B Phys. Biol. Sci., 95, 261–277 (2019).
- 9) Marshansky V, Rubinstein JL, Grüber G. Eukaryotic V-ATPase: Novel structural findings and functional insights. Biochim. Biophys. Acta, 1837, 857–879 (2014).
- 10) Hutton RL, Boyer PD. Subunit interaction during catalysis. Alternating site cooperativity of mitochondrial adenosine triphosphatase. J. Biol. Chem., 254, 9990–9993 (1979).
- 11) Sabbert D, Engelbrecht S, Junge W. Intersubunit rotation in active F-ATPase. Nature, 381, 623–625 (1996).
- 12) Abrahams JP, Leslie AG, Lutter R, Walker JE. Structure at 2.8 Å resolution of F1-ATPase from bovine heart mitochondria. Nature, 370, 621–628 (1994).
- 13) Duncan TM, Bulygin VV, Zhou Y, Hutcheon ML, Cross RL. Rotation of subunits during catalysis by Escherichia coli F1-ATPase. Proc. Natl. Acad. Sci. U.S.A., 92, 10964–10968 (1995).
- 14) Noji H, Yasuda R, Yoshida M, Kinosita K Jr. Direct observation of the rotation of F1-ATPase. Nature, 386, 299–302 (1997).
- 15) Omote H, Sambonmatsu N, Saito K, Sambongi Y, Iwamoto-Kihara A, Yanagida T, Wada Y, Futai M. The γ-subunit rotation and torque generation in F1-ATPase from wild-type or uncoupled mutant Escherichia coli. Proc. Natl. Acad. Sci. U.S.A., 96, 7780–7784 (1999).
- 16) Sambongi Y, Iko Y, Tanabe M, Omote H, Iwamoto-Kihara A, Ueda I, Yanagida T, Wada Y, Futai M. Mechanical rotation of the c subunit oligomer in ATP synthase (F0F1): direct observation. Science, 286, 1722–1724 (1999).
- 17) Tanabe M, Nishio K, Iko Y, Sambongi Y, Iwamoto-Kihara A, Wada Y, Futai M. Rotation of a complex of the γ subunit and c ring of Escherichia coli ATP synthase. The rotor and stator are interchangeable. J. Biol. Chem., 276, 15269–15274 (2001).
- 18) Meiss E, Konno H, Groth G, Hisabori T. Molecular processes of inhibition and stimulation of ATP synthase caused by the phytotoxin tentoxin. J. Biol. Chem., 283, 24594–24599 (2008).
- 19) Nakanishi-Matsui M, Sekiya M, Yano S, Futai M. Inhibition of F1-ATPase rotational catalysis by the carboxyl-terminal domain of the ϵ subunit. J. Biol. Chem., 289, 30822–30831 (2014).
- 20) Sekiya M, Nakamoto RK, Nakanishi-Matsui M, Futai M. Binding of phytopolyphenol piceatannol disrupts β/γ subunit interactions and rate-limiting step of steady-state rotational catalysis in Escherichia coli F1-ATPase. J. Biol. Chem., 287, 22771–22780 (2012).
- 21) Sekiya M, Sakamoto Y, Futai M, Nakanishi-Matsui M. Role of α/β interface in F1 ATPase rotational catalysis probed by inhibitors and mutations. Int. J. Biol. Macromol., 99, 615–621 (2017).
- 22) Baran AA, Silverman KA, Zeskand J, Koratkar R, Palmer A, McCullen K, Curran WJ Jr, Edmonston TB, Siracusa LD, Buchberg AM. The modifier of Min 2 (Mom2) locus: Embryonic lethality of a mutation in the Atp5a1 gene suggests a novel mechanism of polyp suppression. Genome Res., 17, 566–576 (2007).
- 23) Inoue H, Noumi T, Nagata M, Murakami H, Kanazawa H. Targeted disruption of the gene encoding the proteolipid subunit of mouse vacuolar H+-ATPase leads to early embryonic lethality. Biochim. Biophys. Acta., 1413, 130–138 (1999).
- 24) Araki M, Hoshi K, Fujiwara M, Sasaki Y, Yonezawa H, Senpuku H, Iwamoto-Kihara A, Maeda M. Complementation of the Fo c subunit of Escherichia coli with that of Streptococcus mutans and properties of the hybrid FoF1 ATP synthase. J. Bacteriol., 195, 4873–4878 (2013).
- 25) Grosser MR, Paluscio E, Thurlow LR, Dillon MM, Cooper VS, Kawula TH, Richardson AR. Genetic requirements for Staphylococcus aureus nitric oxide resistance and virulence. PLOS Pathog., 14, e1006907 (2018).
- 26) Sasaki M, Kodama Y, Shimoyama Y, Ishikawa T, Kimura S. Aciduricity and acid tolerance mechanisms of Streptococcus anginosus. J. Gen. Appl. Microbiol., 64, 174–179 (2018).
- 27) Sekiya M, Izumisawa S, Iwamoto-Kihara A, Fan Y, Shimoyama Y, Sasaki M, Nakanishi-Matsui M. Proton-pumping F-ATPase plays an important role in Streptococcus mutans under acidic conditions. Arch. Biochem. Biophys., 666, 46–51 (2019).
- 28) Sekiya M, Shimoyama Y, Ishikawa T, Sasaki M, Futai M, Nakanishi-Matsui M. Porphyromonas gingivalis is highly sensitive to inhibitors of a proton-pumping ATPase. Biochem. Biophys. Res. Commun., 498, 837–841 (2018).
- 29) Andries K, Verhasselt P, Guillemont J, Göhlmann HW, Neefs JM, Winkler H, Van Gestel J, Timmerman P, Zhu M, Lee E, Williams P, de Chaffoy D, Huitric E, Hoffner S, Cambau E, Truffot-Pernot C, Lounis N, Jarlier V. A diarylquinoline drug active on the ATP synthase of Mycobacterium tuberculosis. Science, 307, 223–227 (2005).
- 30) Dadi PK, Ahmad M, Ahmad Z. Inhibition of ATPase activity of Escherichia coli ATP synthase by polyphenols. Int. J. Biol. Macromol., 45, 72–79 (2009).
- 31) Hong S, Pedersen PL. ATP synthase and the actions of inhibitors utilized to study its roles in human health, disease, and other scientific areas. Microbiol. Mol. Biol. Rev., 72, 590–641 (2008).
- 32) Sekiya M, Chiba E, Satoh M, Yamakoshi H, Iwabuchi Y, Futai M, Nakanishi-Matsui M. Strong inhibitory effects of curcumin and its demethoxy analog on Escherichia coli ATP synthase F1 sector. Int. J. Biol. Macromol., 70, 241–245 (2014).
- 33) Anand P, Kunnumakkara AB, Newman RA, Aggarwal BB. Bioavailability of curcumin: problems and promises. Mol. Pharm., 4, 807–818 (2007).
- 34) Setoguchi Y, Oritani Y, Ito R, Inagaki H, Maruki-Uchida H, Ichiyanagi T, Ito T. Absorption and metabolism of piceatannol in rats. J. Agric. Food Chem., 62, 2541–2548 (2014).
- 35) Boyer PD. The ATP synthase--a splendid molecular machine. Annu. Rev. Biochem., 66, 717–749 (1997).
- 36) Senior AE, Nadanaciva S, Weber J. The molecular mechanism of ATP synthesis by F1F0-ATP synthase. Biochim. Biophys. Acta BBABioenerg., 1553, 188–211 (2002).
- 37) Stock D, Gibbons C, Arechaga I, Leslie AG, Walker JE. The rotary mechanism of ATP synthase. Curr. Opin. Struct. Biol., 10, 672–679 (2000).
- 38) Walker JE. The ATP synthase: the understood, the uncertain and the unknown. Biochem. Soc. Trans., 41, 1–16 (2013).
- 39) Nakanishi-Matsui M, Kashiwagi S, Ubukata T, Iwamoto-Kihara A, Wada Y, Futai M. Rotational catalysis of Escherichia coli ATP synthase F1 sector. Stochastic fluctuation and a key domain of the β subunit. J. Biol. Chem., 282, 20698–20704 (2007).
- 40) Baylis Scanlon JA, Al-Shawi MK, Le NP, Nakamoto RK. Determination of the partial reactions of rotational catalysis in F1-ATPase. Biochemistry, 46, 8785–8797 (2007).
- 41) Sekiya M, Nakamoto RK, Al-Shawi MK, Nakanishi-Matsui M, Futai M. Temperature dependence of single molecule rotation of the Escherichia coli ATP synthase F1 sector reveals the importance of γ − β subunit interactions in the catalytic dwell. J. Biol. Chem., 284, 22401–22410 (2009).
- 42) Kashiwagi S, Iwamoto-Kihara A, Kojima M, Nonaka T, Futai M, Nakanishi-Matsui M. Effects of mutations in the β subunit hinge domain on ATP synthase F1 sector rotation: interaction between Ser 174 and Ile 163. Biochem. Biophys. Res. Commun., 365, 227–231 (2008).
- 43) Nakanishi-Matsui M, Kashiwagi S, Hosokawa H, Cipriano DJ, Dunn SD, Wada Y, Futai M. Stochastic high-speed rotation of Escherichia coli ATP synthase F1 sector: the ε subunit-sensitive rotation. J. Biol. Chem., 281, 4126–4131 (2006).
- 44) Sekiya M, Hosokawa H, Nakanishi-Matsui M, Al-Shawi MK, Nakamoto RK, Futai M. Single molecule behavior of inhibited and active states of Escherichia coli ATP synthase F1 rotation. J. Biol. Chem., 285, 42058–42067 (2010).
- 45) Bowler MW, Montgomery MG, Leslie AG, Walker JE. Ground state structure of F1-ATPase from bovine heart mitochondria at 1.9 Å resolution. J. Biol. Chem., 282, 14238–14242 (2007).
- 46) Al-Shawi MK, Ketchum CJ, Nakamoto RK. Energy coupling, turnover, and stability of the F0F1 ATP synthase are dependent on the energy of interaction between γ and β subunits. J. Biol. Chem., 272, 2300–2306 (1997).
- 47) Gledhill JR, Montgomery MG, Leslie AG, Walker JE. Mechanism of inhibition of bovine F1-ATPase by resveratrol and related polyphenols. Proc. Natl. Acad. Sci. U.S.A., 104, 13632–13637 (2007).
- 48) Grüber G, Manimekalai MS, Mayer F, Muller V. ATP synthases from archaea: the beauty of a molecular motor. Biochim. Biophys. Acta BBABioenerg., 1837, 940–952 (2014).
- 49) Singharoy A, Chipot C, Ekimoto T, Suzuki K, Ikeguchi M, Yamato I, Murata T. Rotational mechanism model of the bacterial V1 motor based on structural and computational analyses. Front. Physiol., 10, 46 (2019).
- 50) Arai S, Saijo S, Suzuki K, Mizutani K, Kakinuma Y, Ishizuka-Katsura Y, Ohsawa N, Terada T, Shirouzu M, Yokoyama S, Iwata S, Yamato I, Murata T. Rotation mechanism of Enterococcus hirae V1-ATPase based on asymmetric crystal structures. Nature, 493, 703–707 (2013).
- 51) Iida T, Minagawa Y, Ueno H, Kawai F, Murata T, Iino R. Single-molecule analysis reveals rotational substeps and chemo-mechanical coupling scheme of Enterococcus hirae V1-ATPase. J. Biol. Chem., 294, 17017–17030 (2019).
- 52) Imamura H, Nakano M, Noji H, Muneyuki E, Ohkuma S, Yoshida M, Yokoyama K. Evidence for rotation of V1-ATPase. Proc. Natl. Acad. Sci. U.S.A., 100, 2312–2315 (2003).
- 53) Imamura H, Ikeda C, Yoshida M, Yokoyama K. The F subunit of Thermus thermophilus V1-ATPase promotes ATPase activity but is not necessary for rotation. J. Biol. Chem., 279, 18085–18090 (2004).
- 54) Iwabe N, Kuma K, Hasegawa M, Osawa S, Miyata T. Evolutionary relationship of archaebacteria, eubacteria, and eukaryotes inferred from phylogenetic trees of duplicated genes. Proc. Natl. Acad. Sci. U.S.A., 86, 9355–9359 (1989).
- 55) Schäfer IB, Bailer SM, Duser MG, Borsch M, Bernal RA, Stock D, Gruber G. Crystal structure of the archaeal A1Ao ATP synthase subunit B from Methanosarcina mazei Go1: implications of nucleotide-binding differences in the major A1Ao subunits A and B. J. Mol. Biol., 358, 725–740 (2006).
- 56) Foo CS-Y, Pethe K, Lupien A. Oxidative phosphorylation—an update on a new, essential target space for drug discovery in Mycobacterium tuberculosis. Appl. Sci., 10, 2339 (2020).
- 57) Rao SP, Alonso S, Rand L, Dick T, Pethe K. The protonmotive force is required for maintaining ATP homeostasis and viability of hypoxic, nonreplicating Mycobacterium tuberculosis. Proc. Natl. Acad. Sci. U.S.A., 105, 11945–11950 (2008).
- 58) Ahn SJ, Wen ZT, Burne RA. Effects of oxygen on virulence traits of Streptococcus mutans. J. Bacteriol., 189, 8519–8527 (2007).
- 59) García-Godoy F, Hicks MJ. Maintaining the integrity of the enamel surface: the role of dental biofilm, saliva and preventive agents in enamel demineralization and remineralization. J. Am. Dent. Assoc., 139 (Suppl. 2), 25S–34S (2008).
- 60) Matsui R, Cvitkovitch D. Acid tolerance mechanisms utilized by Streptococcus mutans. Future Microbiol., 5, 403–417 (2010).
- 61) Kuhnert WL, Zheng G, Faustoferri RC, Quivey RG Jr. The F-ATPase operon promoter of Streptococcus mutans is transcriptionally regulated in response to external pH. J. Bacteriol., 186, 8524–8528 (2004).
- 62) Rahman M, Nguyen SV, McCullor KA, King CJ, Jorgensen JH, McShan WM. Comparative genome analysis of the daptomycin-resistant Streptococcus anginosus strain J4206 associated with breakthrough bacteremia. Genome Biol. Evol., 8, 3446–3459 (2016).
- 63) Terzi HA, Demiray T, Koroglu M, Cakmak G, Hakki Ciftci I, Ozbek A, Altindis M. Intra-abdominal abscess and primary peritonitis caused by Streptococcus anginosus. Jundishapur J. Microbiol., 9, e33863 (2016).
- 64) Willcox MD. Potential pathogenic properties of members of the “Streptococcus milleri” group in relation to the production of endocarditis and abscesses. J. Med. Microbiol., 43, 405–410 (1995).
- 65) Kawasaki M, Ikeda Y, Ikeda E, Takahashi M, Tanaka D, Nakajima Y, Arakawa S, Izumi Y, Miyake S. Oral infectious bacteria in dental plaque and saliva as risk factors in patients with esophageal cancer. Cancer, 127, 512–519 (2021).
- 66) Sasaki H, Igaki H, Ishizuka T, Kogoma Y, Sugimura T, Terada M. Presence of Streptococcus DNA sequence in surgical specimens of gastric cancer. Jpn. J. Cancer Res., 86, 791–794 (1995).
- 67) Sasaki M, Yamaura C, Ohara-Nemoto Y, Tajika S, Kodama Y, Ohya T, Harada R, Kimura S. Streptococcus anginosus infection in oral cancer and its infection route. Oral Dis., 11, 151–156 (2005).
- 68) Belli WA, Marquis RE. Adaptation of Streptococcus mutans and Enterococcus hirae to acid stress in continuous culture. Appl. Environ. Microbiol., 57, 1134–1138 (1991).
- 69) Slots J, Listgarten MA. Bacteroides gingivalis, Bacteroides intermedius and Actinobacillus actinomycetemcomitans in human periodontal diseases. J. Clin. Periodontol., 15, 85–93 (1988).
- 70) Könönen E, Conrads G, Nagy E. Man. Clin. Microbiol., 967-93 (2015).
- 71) Ohara-Nemoto Y, Sarwar MT, Shimoyama Y, Kobayakawa T, Nemoto TK. Preferential dipeptide incorporation of Porphyromonas gingivalis mediated by proton-dependent oligopeptide transporter (Pot). FEMS Microbiol. Lett., 367, 1–8 (2020).
- 72) Flanagan D. Enterococcus faecalis and dental Implants. J. Oral Implantol., 43, 8–11 (2017).
- 73) Pinheiro ET, Mayer MPA. Enterococcus faecalis in oral infections. JBR J. Interdiscipl. Med. Dent. Sci., 3, 160 (2015).
- 74) Kakinuma Y, Igarashi K. Mutants of Streptococcus faecalis sensitive to alkaline pH lack Na+-ATPase. J. Bacteriol., 172, 1732–1735 (1990).
- 75) Hermolin J, Fillingame RH. H+-ATPase activity of Escherichia coli F1F0 is blocked after reaction of dicyclohexylcarbodiimide with a single proteolipid (subunit c) of the F0 complex. J. Biol. Chem., 264, 3896–3903 (1989).
- 76) Ivey DM, Ljungdahl LG. Purification and characterization of the F1-ATPase from Clostridium thermoaceticum. J. Bacteriol., 165, 252–257 (1986).
- 77) Satre M, Bof M, Vignais PV. Interaction of Escherichia coli adenosine triphosphatase with aurovertin and citreoviridin: Inhibition and fluorescence studies. J. Bacteriol., 142, 768–776 (1980).
- 78) Lemker T, Ruppert C, Stoger H, Wimmers S, Muller V. Overproduction of a functional A1 ATPase from the archaeon Methanosarcina mazei Go1 in Escherichia coli. Eur. J. Biochem., 268, 3744–3750 (2001).
- 79) Guo H, Courbon GM, Bueler SA, Mai J, Liu J, Rubinstein JL. Structure of mycobacterial ATP synthase bound to the tuberculosis drug bedaquiline. Nature, 589, 143–147 (2021).
- 80) Hards K, McMillan DGG, Schurig-Briccio LA, Gennis RB, Lill H, Bald D, Cook GM. Ionophoric effects of the antitubercular drug bedaquiline. Proc. Natl. Acad. Sci. U.S.A., 115, 7326–7331 (2018).
- 81) Koul A, Vranckx L, Dendouga N, Balemans W, Van den Wyngaert I, Vergauwen K, Göhlmann HWH, Willebrords R, Poncelet A, Guillemont J, Bald D, Andries K. Diarylquinolines are bactericidal for dormant mycobacteria as a result of disturbed ATP homeostasis. J. Biol. Chem., 283, 25273–25280 (2008).
- 82) WHO. Global tuberculosis Report. (2021).
- 83) Lamontagne Boulet M, Isabelle C, Guay I, Brouillette E, Langlois JP, Jacques PÉ, Rodrigue S, Brzezinski R, Beauregard PB, Bouarab K, Boyapelly K, Boudreault PL, Marsault É, Malouin F. Tomatidine Is a Lead Antibiotic Molecule That Targets Staphylococcus aureus ATP Synthase Subunit C. Antimicrob. Agents Chemother., 62, e02197-17 (2018).
- 84) Wu C, Luo J, Wu M, Meng F, Cai Z, Chen Y, Sun T. Design, synthesis and biological evaluation of anti-tuberculosis agents based on bedaquiline structure. Med. Chem., 16, 703–714 (2020).
- 85) Tsao R. Chemistry and biochemistry of dietary polyphenols. Nutrients, 2, 1231–1246 (2010).
- 86) Ahmad Z, Hassan SS, Azim S. A therapeutic connection between dietary phytochemicals and ATP synthase. Curr. Med. Chem., 24, 3894–3906 (2017).
- 87) Ahmad Z, Laughlin TF, Kady IO. Thymoquinone inhibits Escherichia coli ATP synthase and cell growth. PLOS ONE, 10, e0127802 (2015).
- 88) Amini A, Liu M, Ahmad Z. Understanding the link between antimicrobial properties of dietary olive phenolics and bacterial ATP synthase. Int. J. Biol. Macromol., 101, 153–164 (2017).
- 89) Sekiya M, Hisasaka R, Iwamoto-Kihara A, Futai M, Nakanishi-Matsui M. A unique mechanism of curcumin inhibition on F1 ATPase. Biochem. Biophys. Res. Commun., 452, 940–944 (2014).
- 90) Zheng J, Ramirez VD. Inhibition of mitochondrial proton F0F1-ATPase/ATP synthase by polyphenolic phytochemicals. Br. J. Pharmacol., 130, 1115–1123 (2000).