Abstract
Pyridoxine (VB6) is a vitamin that is essential to maintain the homeostasis of the human body by contributing to various metabolic reactions. In the skin, although some studies have shown that VB6 is involved in regulating homeostasis through the attenuation of intracellular oxidative stress, there are few reports regarding the effects of VB6 on the prevention or improvement of skin aging. Thus, we conducted this study to determine the potential anti-skin pigmentation effect of VB6 focusing on the phagocytosis of melanosomes (MSs) by keratinocytes. The phagocytosis of MSs by keratinocytes is activated by oxidative stress and is an important factor of skin pigmentation and the eventual appearance of pigmented spots. First, we confirmed the antioxidant property of VB6 that enhanced the expression of several intracellular antioxidants via nuclear erythroid factor 2-related factor 2 (Nrf2). Although the incorporation of fluorescent beads (FBs), which are used as pseudo-MSs, into keratinocytes was increased under higher oxidation conditions caused by UVB and by the depletion of intracellular glutathione, treatment with VB6 suppressed the increased incorporation of FBs into those keratinocytes via Nrf2 activation. Furthermore, VB6 restored the decreased expression of differentiation marker proteins in keratinocytes caused by FB incorporation. Taken together, the results show that VB6 has the potential to prevent the appearance of pigmented spots by suppressing the activation of phagocytosis in keratinocytes caused by oxidative stress, and by restoring the differentiation of keratinocytes disrupted by FB incorporation.
INTRODUCTION
Pyridoxine (VB6), which is a water-soluble vitamin, is known to act as a coenzyme for enzymes involved in amino acid metabolism and nutrition, and which contributes to maintain the homeostasis of the body.1,2) In the skin, the depletion of VB6 causes some disorders like atopic dermatitis and seborrheic dermatitis.3,4) These facts suggest that VB6 is necessary to maintain homeostasis in the skin.
Meanwhile, the skin is always exposed to oxidative stress because of its location at the surface of the human body. External circumstances, such as sunlight, air pollution, high temperature and low humidity, stimulate the generation of reactive oxygen species (ROS).5–8) Among those stresses, the UV radiation in sunlight generates ROS excessively through the elevation of calcium influx in keratinocytes and results in accelerating the appearance of skin aging such as wrinkles, sagging and pigmented spots, like solar lentigos.9–12) Among the features of skin aging, faces that have many pigmented spots give a strong impression of aging and may reduce their social activities. Thus, to prevent or improve the appearance of pigmented spots is an important challenge to rescue the decreased social activity for all human society.
Pigmented spots are recognized as limited areas of skin hyperpigmentation. Although the reasons for the appearance of pigmentation in limited areas is still unknown, the mechanisms involved in the formation of pigmented spots is similar to the mechanism regulating skin pigmentation.12) In general, although skin pigmentation occurs due to UV exposure or inflammation, increases of melanin synthesis by the activation of melanocytes are essential in pigmentation.13) Additionally, the diffusion of melanosomes (MSs), in which melanin is produced and stored in melanocytes, into the epidermis due to their incorporation into keratinocytes by phagocytosis has been recognized as one determinant of skin color and also is a critical process of skin pigmentation.14) Recently, oxidative stress was shown to enhance the phagocytosis of MSs into keratinocytes.15,16) In fact, UVB, which is a strong generator of ROS, enhances the phagocytosis of MSs isolated from melanocytes and of fluorescent beads (FBs), which are used as pseudo-MSs.16,17) MSs incorporated into keratinocytes by phagocytosis are exfoliated from the skin surface by desquamation according to the differentiation of keratinocytes.18) Thus, the prevention or improvement of pigmented spots can be achieved not only by inactivating melanocytes and melanin synthesis but also by suppressing the phagocytosis of MSs and accelerating the terminal differentiation of keratinocytes to exfoliate MSs from the skin.
Interestingly, it has been revealed that VB6 has the ability to attenuate intracellular oxidative stress not only by directly scavenging ROS, but also by enhancing glutathione (GSH) synthesis through the activation of nuclear erythroid factor 2 (NE-F2)-related factor 2 (Nrf2) in keratinocytes.19,20) The binding of Nrf2 to an antioxidant response element after translocation from the cytoplasm into the nucleus up-regulates multiple types of antioxidant-related genes, including γ-glutamylcysteine synthetase (γ-GCS), which is the rate-limiting enzyme of GSH synthesis, catalase, heme oxigenase-1 (HO-1) and nicotinamide adenine dinucleotide phosphate (NADPH) quinone oxidoreductase 1 (NQO1).21,22) Thus, these considerations suggest that VB6 is an important antioxidant to keep the redox balance in keratinocytes, and prompted us to characterize the potential of VB6 as an anti-skin aging substance focusing on the prevention or improvement of pigmented spots.
In this study, we evaluated the effects of VB6 focusing on the phagocytosis of MSs into keratinocytes and its influence on the differentiation of keratinocytes in order to investigate the potential of VB6 to have an anti-skin pigmentation.
MATERIALS AND METHODS
ReagentsHuMedia-KG2 was purchased from Kurabo Industries (Osaka, Japan). Buthionine sulfoximine (BSO) and pyridoxine hydrochloride were purchased from Nacalai Tesque (Kyoto, Japan). Fluorescent beads (FluoSpheres® carboxylate-modified 0.2 µm), the BCA protein assay kit, Nrf2-small interfering RNA (siRNA), non-targeting siRNA, Lipofectamine RNAiMAX, the Power SYBR Green Cells-to-CT kit and Mammalian Protein Extraction Reagent were purchased from Thermo Fisher Scientific (Waltham, MA, U.S.A.). Hoechst 33342 was purchased from Dojindo (Kumamoto, Japan). The anti-keratin 10 polyclonal antibody and the anti-actin polyclonal antibody were purchased from Abcam (Cambridge, U.K.). The anti-involucrin polyclonal antibody and the anti-loricrin polyclonal antibody were purchased from Proteintech (Rosemont, IL, U.S.A.). EzWestBlue was purchased from ATTO (Tokyo, Japan).
Cell CultureNormal human epidermal keratinocytes (NHEKs) derived from the foreskins of Asian and Caucasian donors were obtained from Kurabo (Osaka, Japan) and were cultured in HuMedia-KG2 at 37 °C in a 5% CO2 atmosphere.
RNA InterferenceNHEKs were seeded into multi-well plates in HuMedia-KG2, and were treated with 100 nM Nrf2-siRNA (siNrf2) or non-targeting siRNA (siControl) using Lipofectamine RNAiMAX for 24 h.
Real-Time PCRTotal RNAs were extracted from NHEKs and cDNAs were synthesized using a Power SYBR Green Cells-to-CT kit. Real-time PCR was performed with SYBR Green Master Mix using an Eco Real-Time PCR System (Illumina, San Diego, CA, U.S.A.). Data were analyzed according to the ΔΔCt method.
Incorporation of FBs into KeratinocytesAfter treatment of NHEKs, 100 µM BSO pretreated NHEKs or siNrf2-treated NHEKs with samples for 24 h, the cells were irradiated with UVB at 20 mJ/cm2. At 4 h after incubation with FBs,16) the incorporation of FBs into cells was observed using a Floid® Cell Imaging Station, and was quantified by the measurement of fluorescence intensity (Ex; 535 nm, Em; 590 nm). The UVB source used was a TL20W/12RS UVB broadband lamp (Philips, Eindhoven, Holland, Netherlands) and radiation energies were measured with a UVX radiometer (UVP, Upland, Canada). In addition, protein concentrations were determined using a BCA protein assay kit.
Western BlottingCellular proteins were extracted with a protein extraction reagent containing protease inhibitors. The extracted proteins were separated by sodium dodecyl sulfate (SDS)-polyacrylamide gel electrophoresis (PAGE) on 10% polyacrylamide gels, and then were transferred to polyvinylidene difluoride membranes. The membranes were then incubated with primary antibodies specific for keratin 10 (1 : 1000), involucrin (1 : 1000), loricrin (1 : 1000) and β-actin (1 : 2000). Immunoreactive protein bands were then visualized using EzWestBlue and were analyzed using Image J.
Statistical AnalysisAll data are expressed as means ± standard deviation (S.D.). Comparisons between two groups or multiple groups were performed by Student’s t-test or Tukey’s test respectively. A p-value <0.05 is considered statistically significant.
RESULTS
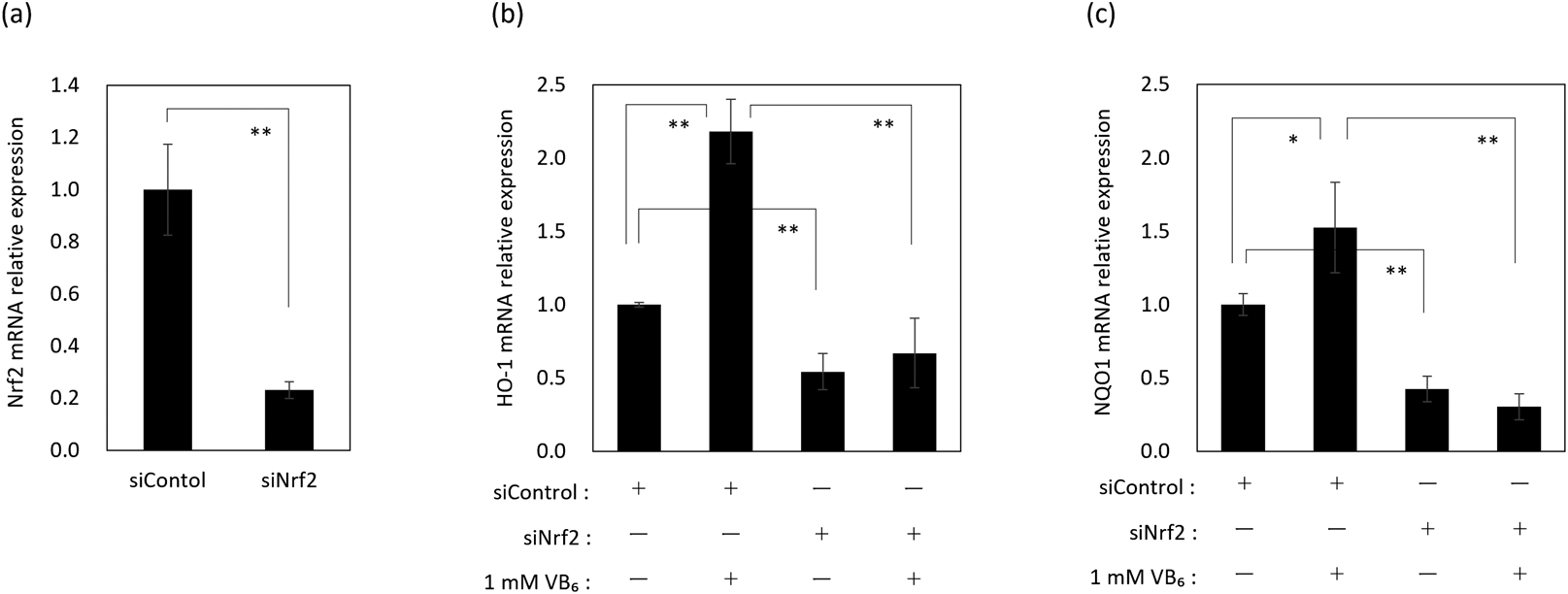
Identification of the Intracellular Antioxidant Ability of VB6Our previous study revealed that VB6 stimulated the nuclear translocation of Nrf2 and up-regulated the mRNA expression level of γ-GCS in keratinocytes.20) Here, in order to confirm in detail the antioxidant actions of VB6 in keratinocytes and the involvement of Nrf2, mRNA expression levels of other antioxidants, such as HO-1 and NQO1 that are encoded downstream of Nrf2, in VB6 treated NHEKs were examined using a siNrf2 knockdown system (Fig. 1a). mRNA expression levels of HO-1 and NQO1 were significantly up-regulated at 24 h after treatment with VB6 (Figs. 1b, c). However, the up-regulation of those mRNA expression levels by VB6 was abolished by Nrf2 knockdown in NHEKs (Figs. 1b, c). These results indicated that VB6 enhanced the intracellular antioxidant ability due to the up-regulation of mRNA expression levels of HO-1 and NQO1 via Nrf2.
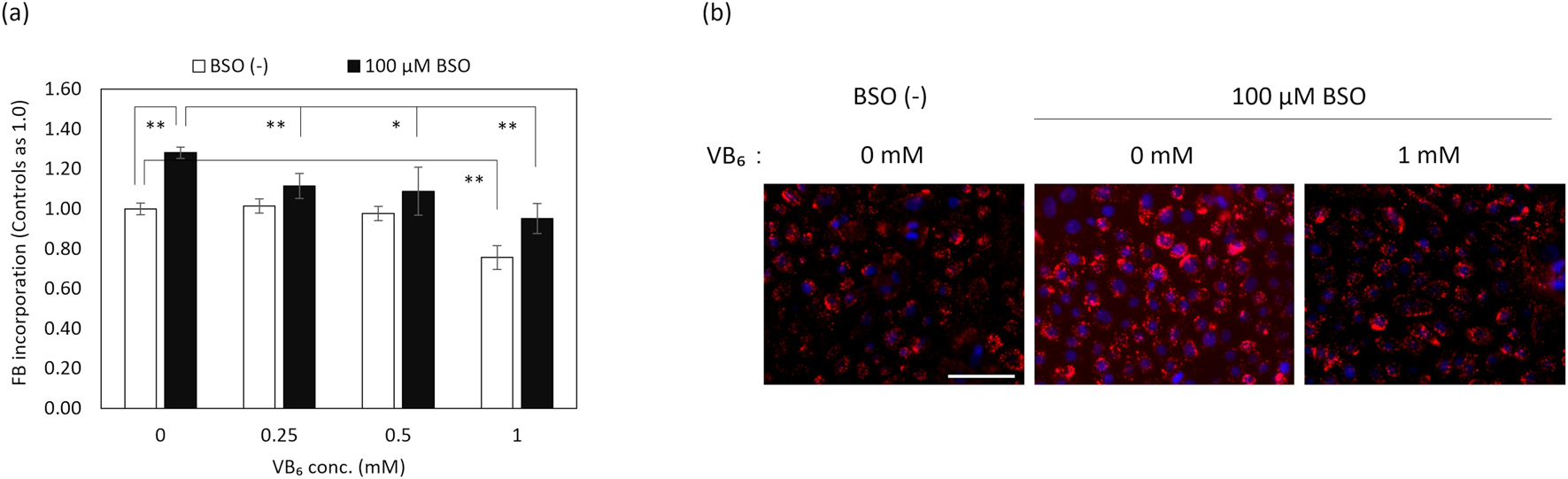
Influence of VB6 on the Incorporation of FBs in NHEKs in Higher Oxidative SituationsIn order to evaluate the effects of VB6 on the phagocytic ability of keratinocytes in higher oxidative situations, we examined the ratio of FB incorporation into NHEKs. NHEKs in higher oxidative situations without cytotoxicity were prepared by the depletion of GSH due to the inhibition of GSH synthesis with BSO,23) and by irradiation with UVB. Treatment with VB6 caused a significant suppression of the incorporation of FBs in NHEKs elevated by UVB exposure and by BSO treatment (Figs. 2, 3).
Mechanism by Which VB6 Suppresses FB Incorporation Elevated by UVB in NHEKsWe then examined the involvement of Nrf2 in the reduced FB incorporation into NHEKs caused by VB6 using Nrf2 knockdown. Interestingly, the suppression of FB incorporation caused by VB6 in UVB-irradiated NHEKs was abrogated by the treatment with siNrf2 (Fig. 4).
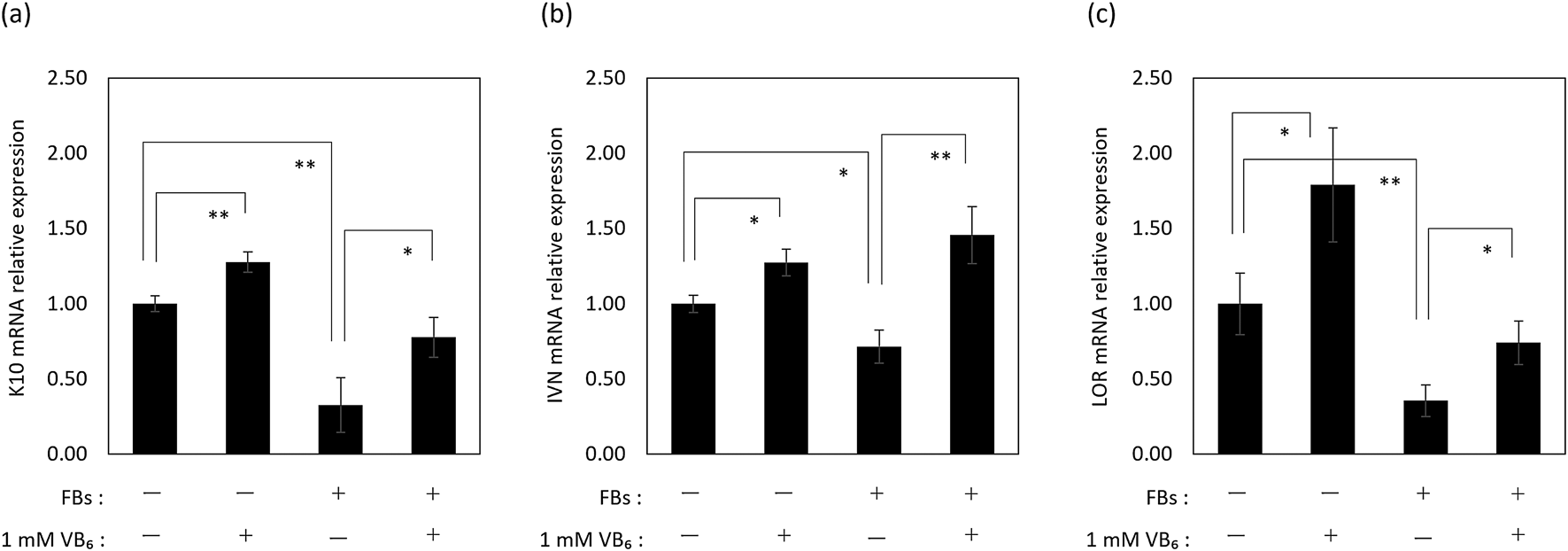
Effects of VB6 on the Differentiation in FB-Incorporated NHEKsIt has been reported that the epidermis in pigmented areas of solar lentigos due to the high incorporation of MSs shows decreased expression of keratin 10 (K10), involucrin (IVN) and loricrin (LOR), which are differentiation markers of keratinocytes.24,25) This fact suggests that the differentiation of keratinocytes is disrupted by the incorporation of materials like MSs. So, we examined the influence of FB incorporation on the differentiation of NHEKs at 1 or 5 d after FB incorporation.
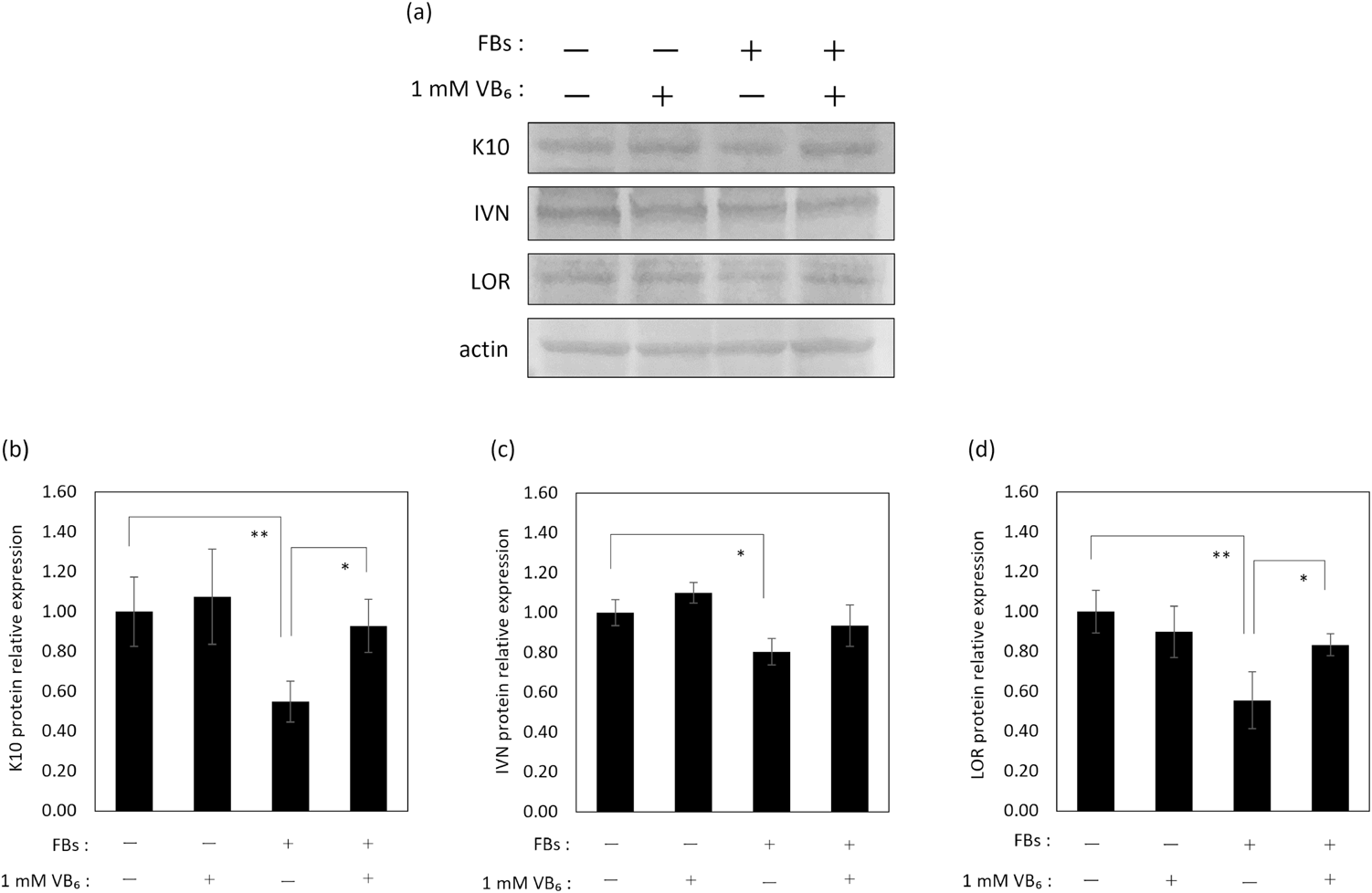
The results show that the mRNA expression levels of K10, IVN and LOR were down-regulated in NHEKs that had incorporated FBs 1 d earlier (Fig. 5). Additionally, those protein expression levels were confirmed to be decreased at 5 d after FB incorporation (Fig. 6). These results confirmed experimentally that the incorporation of materials such as FBs disrupted the differentiation of keratinocytes at the mRNA and protein expression levels.
Treatment with VB6 significantly restored the mRNA expression level of those differentiation markers at 1 d after FB incorporation (Fig. 5). Further, a significant restoration of K10 and LOR protein expression levels in NHEKs was found at 5 d after FB incorporation (Figs. 6a, b, d). Regarding the protein expression level of IVN, its restoration by VB6 was confirmed (Figs. 6a, c). However, VB6 did not show any influence on the expression of differentiation markers in FB-unincorporated NHEKs (Fig. 6).
DISCUSSION
Pigmented spots like solar lentigos are a type of skin hyperpigmentation that appears at small limited areas and their appearance is accelerated by chronic sun exposure.12,13,24) However, the reasons for the appearance of hyperpigmentation in limited areas is still unknown. To date, it has been well demonstrated that ROS is a major trigger of UV-induced skin pigmentation. ROS generated in keratinocytes enhances melanin synthesis in MSs by melanocytes through the binding of melanocyte activating substances such as α-melanocyte-stimulating hormone, stem cell factor and endothelin-1 (ET-1) secreted from keratinocytes that bind to specific receptors on melanocytes (melanocortin 1 receptor/c-kit/ET-BR).13,26) Mature MSs containing abundant melanin are transported from the perinuclear area of melanocytes to the tips of dendrites and finally they are incorporated into neighboring keratinocytes by phagocytosis.27–30) Thus, skin pigmentation results not only due to increases of melanin synthesis but also due to the diffusion of MSs by phagocytosis into keratinocytes. In a previous study, we reported that external or internal oxidative stresses caused by hydrogen peroxide or by the depletion of glutathione by BSO enhanced the phagocytic ability of keratinocytes.16) Thus, the sum of these results suggests that a higher oxidative environment in keratinocytes is one of the key events that initiates UV-induced pigmentation.
On the other hand, although previous studies reported that VB6 has the potential to improve skin problems due to its antioxidant property,19,20) the anti-aging effects of VB6 in the skin has not been fully demonstrated. Here, to demonstrate the potential of VB6 to prevent skin pigmentation, we examined the effects of VB6 on the incorporation of MSs into keratinocytes by phagocytosis in a higher oxidative environment due to UVB irradiation.
We first confirmed the antioxidant profile of VB6, focusing on the expression of several antioxidant enzymes, HO-1 and NQO1, that are encoded downstream of Nrf2. VB6 enhanced the mRNA expression levels of HO-1 and NQO1 via Nrf2 as well as γ-GCS, as previously we reported20) (Fig. 1). These results confirmed that VB6 in fact enhances the anti-oxidative defense in keratinocytes via the activation of Nrf2.
UVB and GSH depletion by BSO increased intracellular ROS levels in NHEKs (data not shown). Next, we found that VB6 suppressed the incorporation of FBs into NHEKs under higher oxidative situations caused by UVB or GSH depletion (Figs. 2–4). Additionally, the suppression of VB6 on the elevated incorporation of FBs was canceled by the knockdown of Nrf2 (Fig. 4), which suggested that VB6 suppresses FB incorporation elevated by UVB in NHEKs through the amelioration of intracellular oxidative stress via Nrf2.
In this study, we used FBs instead of MSs to evaluate the incorporation of MSs into keratinocytes by phagocytosis. Because keratinocytes which have incorporated FBs show similar behaviors to keratinocytes that have incorporated MSs, FBs are often used instead of MSs in studies of MS incorporation into keratinocytes.16,17,31–34) Furthermore, the use of FBs instead of MSs makes it possible to evaluate phagocytosis quantitatively and accurately.16,17,31–34) Summarizing these facts, we deduced that VB6 suppresses the incorporation of MSs into keratinocytes in higher oxidative situations.
Considering the fate of MSs incorporated into keratinocytes, MSs are exfoliated from the skin surface through the desquamation of corneocytes, which are the end-products of the differentiation of keratinocytes.18) It is known that the characteristics of the epidermis in pigmented areas are different from non-pigmented epidermis at peripheral regions. One notable characteristic is that the rates of proliferation and of differentiation of keratinocytes in pigmented areas are slower than keratinocytes in non-pigmented peripheral areas.24,25,35) In fact, MSs incorporated into keratinocytes in the basal layer do not express Ki67, which is a proliferation marker,36) and additionally pigmented epidermis shows a lower expression of differentiation marker proteins.24,25) These facts suggest that the incorporation of MSs is one cause that disturbs epidermal homeostasis, which is an imbalance of supplying progenitor keratinocytes and differentiated keratinocytes by decreased proliferation and differentiation, resulting in a long-term retention of MSs in the epidermis by suppressing the exfoliation of corneocytes from the surface. In this study, we found that NHEKs that incorporated FBs showed lower expression levels of K10, IVN and LOR, which are early, middle and late markers of differentiation, respectively, compared with NHEKs without FBs (Figs. 5, 6). Thus, we consider that the differentiation of keratinocytes is suppressed by incorporating materials like MSs and FBs. Meanwhile, pretreatment with VB6 restored the decreases of mRNA and protein expression levels of those differentiation markers in NHEKs incorporated with FBs, despite the fact that VB6 didn’t affect the protein expression levels of those differentiation markers in NHEKs without FBs (Figs. 5, 6). These results indicate that VB6 specifically improves the decreased differentiation of NHEKs caused by the incorporation of FBs. Currently, the mechanism involved in the suppression of differentiation marker proteins by the incorporation of FBs or MSs is unclear. It is interesting that VB6 improved the disruption of differentiation caused by the incorporation of FBs or MSs. The mechanism by which VB6 improves the differentiation of keratinocytes, including the mechanism where differentiation is disturbed by the incorporation of materials into keratinocytes requires future study.
Taken together, our study demonstrated that VB6 suppresses the activation of the phagocytic behavior of keratinocytes in higher oxidative situations by increasing the expression levels of antioxidant enzymes via the activation of Nrf2. Additionally, VB6 improved the decreased differentiation of keratinocytes caused by the incorporation of FBs. These experimental results support the possibility that VB6 will prevent the appearance of pigmented spots due to increasing levels of intracellular antioxidants and improving the differentiation of keratinocytes caused by the incorporation of MSs. We thus conclude that VB6 has the potential to be an anti-skin pigmentation substance.
Conflict of Interest
This study was funded by a research Grant provided by the SSP Co., Ltd. Koichi Hiyama, Atsushi Sawamura and Ichiro Kawase are employees of SSP Co., Ltd. All authors do not have shares of SSP Co., Ltd.
REFERENCES
- 1) Mason JB, Miller JW. The effects of vitamins B12, B6, and folate on blood homocysteine levels. Ann. N. Y. Acad. Sci., 669, 197–203, discussion, 203–204 (1992).
- 2) Rosenberg IH. A history of the isolation and identification of vitamin B(6). Ann. Nutr. Metab., 61, 236–238 (2012).
- 3) Seguchi T, Cui CY, Kusuda S, Takahashi M, Aisu K, Tezuka T. Decreased expression of filaggrin in atopic skin. Arch. Dermatol. Res., 288, 442–446 (1996).
- 4) Schereiner AW, Slinger W, Hawkins VR, Vilter RW. Seborrheic dermatitis; a local metabolic defect involving pyridoxine. J. Lab. Clin. Med., 40, 121–130 (1952).
- 5) Polefka TG, Meyer TA, Agin PP, Bianchini RJ. Effects of solar radiation on the skin. J. Cosmet. Dermatol., 11, 134–143 (2012).
- 6) Jin SP, Li Z, Choi EK, Lee S, Kim YK, Seo EY, Chung JH, Cho S. Urban particulate matter in air pollution penetrates into the barrier-disrupted skin and produces ROS-dependent cutaneous inflammatory response in vivo. J. Dermatol. Sci., 91, 175–183 (2018).
- 7) Cho S, Shin MH, Kim YK, Seo JE, Lee YM, Park CH, Chung JH. Effects of infrared radiation and heat on human skin aging in vivo. J. Investig. Dermatol. Symp. Proc., 14, 15–19 (2009).
- 8) Yokota M, Shimizu K, Kyotani D, Yahagi S, Hashimoto S, Masaki H. The possible involvement of skin dryness on alterations of the dermal matrix. Exp. Dermatol., 23 (Suppl. 1), 27–31 (2014).
- 9) Masaki H, Izutsu Y, Yahagi S, Okano Y. Reactive oxygen species in HaCaT keratinocytes after UVB irradiation are triggered by intracellular Ca(2+) levels. J. Investig. Dermatol. Symp. Proc., 14, 50–52 (2009).
- 10) Wlaschek M, Tantcheva-Poór I, Naderi L, Ma W, Schneider LA, Razi-Wolf Z, Schüller J, Scharffetter-Kochanek K. Solar UV irradiation and dermal photoaging. J. Photochem. Photobiol. B, 63, 41–51 (2001).
- 11) Imokawa G, Ishida K. Biological mechanisms underlying the ultraviolet radiation-induced formation of skin wrinkling and sagging I: reduced skin elasticity, highly associated with enhanced dermal elastase activity, triggers wrinkling and sagging. Int. J. Mol. Sci., 16, 7753–7775 (2015).
- 12) Praetorius C, Sturm RA, Steingrimsson E. Sun-induced freckling: ephelides and solar lentigines. Pigment Cell Melanoma Res., 27, 339–350 (2014).
- 13) Gilchrest BA, Park HY, Eller MS, Yaar M. Mechanisms of ultraviolet light-induced pigmentation. Photochem. Photobiol., 63, 1–10 (1996).
- 14) Babiarz-Magee L, Chen N, Seiberg M, Lin CB. The expression and activation of protease-activated receptor-2 correlate with skin color. Pigment Cell Res., 17, 241–251 (2004).
- 15) Li J, Tang LY, Fu WW, Yuan J, Sheng YY, Yang QP. Low-concentration hydrogen peroxide can upregulate keratinocyte intracellular calcium and PAR-2 expression in a human keratinocyte-melanocyte co-culture system. Arch. Dermatol. Res., 308, 723–731 (2016).
- 16) Katsuyama Y, Sato Y, Okano Y, Masaki H. Intracellular oxidative stress induced by calcium influx initiates the activation of phagocytosis in keratinocytes accumulating at S-phase of the cell cycle after UVB irradiation. J. Dermatol. Sci., 103, 41–48 (2021).
- 17) Virador VM, Muller J, Wu X, Abdel-Malek ZA, Yu ZX, Ferrans VJ, Kobayashi N, Wakamatsu K, Ito S, Hammer JA, Hearing VJ. Influence of alpha-melanocyte-stimulating hormone and ultraviolet radiation on the transfer of melanosomes to keratinocytes. FASEB J., 16, 105–107 (2002).
- 18) Wolff K. Melanocyte-keratinocyte interactions in vivo: the fate of melanosomes. Yale J. Biol. Med., 46, 384–396 (1973).
- 19) Bilski P, Li MY, Ehrenshaft M, Daub ME, Chignell CF. Vitamin B6 (pyridoxine) and its derivatives are efficient singlet oxygen quenchers and potential fungal antioxidants. Photochem. Photobiol., 71, 129–134 (2000).
- 20) Mizutani T, Yamawaki Y, Okano Y, Masaki H. Pyridoxine (VB6) restores the downregulation of serine palmitoyltransferase mRNA expression in keratinocytes cultured in highly oxidative conditions through enhancement of the intracellular antioxidant system. Exp. Dermatol., 28 (Suppl. 1), 43–49 (2019).
- 21) Zhang DD. Mechanistic studies of the Nrf2-Keap1 signaling pathway. Drug Metab. Rev., 38, 769–789 (2006).
- 22) Nguyen T, Nioi P, Pickett CB. The Nrf2-antioxidant response element signaling pathway and its activation by oxidative stress. J. Biol. Chem., 284, 13291–13295 (2009).
- 23) Griffith OW, Meister A. Potent and specific inhibition of glutathione synthesis by buthionine sulfoximine (S-n-butyl homocysteine sulfoximine). J. Biol. Chem., 254, 7558–7560 (1979).
- 24) Choi W, Yin L, Smuda C, Batzer J, Hearing VJ, Kolbe L. Molecular and histological characterization of age spots. Exp. Dermatol., 26, 242–248 (2017).
- 25) Goorochurn R, Viennet C, Granger C, Fanian F, Varin-Blank N, Roy CL, Humbert P. Biological processes in solar lentigo: insights brought by experimental models. Exp. Dermatol., 25, 174–177 (2016).
- 26) Videira IFS, Moura DFL, Magina S. Mechanisms regulating melanogenesis. An. Bras. Dermatol., 88, 76–83 (2013).
- 27) Hume AN, Seabra MC. Melanosomes on the move: a model to understand organelle dynamics. Biochem. Soc. Trans., 39, 1191–1196 (2011).
- 28) Hara M, Yaar M, Byers HR, Goukassian D, Fine RE, Gonsalves J, Gilchrest BA. Kinesin participates in melanosomal movement along melanocyte dendrites. J. Invest. Dermatol., 114, 438–443 (2000).
- 29) Van Gele M, Geusens B, Schmitt AM, Aguilar L, Lambert J. Knockdown of myosin Va isoforms by RNAi as a tool to block melanosome transport in primary human melanocytes. J. Invest. Dermatol., 128, 2474–2484 (2008).
- 30) Ando H, Niki Y, Ito M, Akiyama K, Matsui MS, Yarosh DB, Ichihashi M. Melanosomes are transferred from melanocytes to keratinocytes through the processes of packaging, release, uptake, and dispersion. J. Invest. Dermatol., 132, 1222–1229 (2012).
- 31) Cardinali G, Ceccarelli S, Kovacs D, Aspite N, Lotti LV, Torrisi MR, Picardo M. Keratinocyte growth factor promotes melanosome transfer to keratinocytes. J. Invest. Dermatol., 125, 1190–1199 (2005).
- 32) Cardinali G, Bolasco G, Aspite N, Lucania G, Lotti LV, Torrisi MR, Picardo M. Melanosome transfer promoted by keratinocyte growth factor in light and dark skin-derived keratinocytes. J. Invest. Dermatol., 128, 558–567 (2008).
- 33) Belleudi F, Purpura V, Scrofani C, Persechino F, Leone L, Torrisi MR. Expression and signaling of the tyrosine kinase FGFR2b/KGFR regulates phagocytosis and melanosome uptake in human keratinocytes. FASEB J., 25, 170–181 (2011).
- 34) Endo K, Mizutani T, Okano Y, Masaki H. A red pumpkin seed extract reduces melanosome transfer to keratinocytes by activation of Nrf2 signaling. J. Cosmet. Dermatol., 18, 827–834 (2019).
- 35) Choi HI, Sohn KC, Hong DK, Lee Y, Kim CD, Yoon TJ, Park JW, Jung S, Lee JH, Lee YH. Melanosome uptake is associated with the proliferation and differentiation of keratinocytes. Arch. Dermatol. Res., 306, 59–66 (2014).
- 36) Yan X, Wang TM, Ming YC, Yeh YM, Chen TY, Pang JHS. Melanin uptake reduces cell proliferation of human epidermal keratinocytes. J. Cosmet. Dermatol. Sci. Application, 5, 300–310 (2015).