Abstract
Breast cancer (BC) is the most generally diagnosed cancer and the driving cause of cancer-related death. Transmembrane (TMEM) proteins have been reported to serve as prognostic indicators in a variety of cancers, and it can offer therapeutic targets for carcinoma. However, the function of TMEM in BC remains unclear. In this study, TMEM9A, a member of TMEM family, was screened as the candidate gene after analyzing the profiles of GSE42568 and GEPIA-BRCA database via bioinformatic method. The upregulated expression of TMEM9A was confirmed in BC samples compared with the paired normal tissues. Hence, we speculated that TMEM9A might promote BC progression. To test the hypothesis, we performed a series of loss/gain-of-function experiments and found that BC cells with TMEM9A deletion inhibited cell proliferation, migration, and invasion along with induced apoptosis. Conversely, TMEM9A overexpression reversed the trend. Mechanically, TMEM9A knockdown blocked the Wnt/β-catenin signaling pathway as evidenced by the increased adenomatous polyposis coli (APC) expression and decreased β-catenin, cyclin D1, and axis inhibition protein 2 (AXIN2) expression. Furthermore, over-activation of the Wnt/β-catenin pathway by transfecting BC cells with β-Catenin-S33Y (β-Catenin tyrosine for serine at codon 33) plasmids reversed the effects caused by TMEM9A knockdown. In conclusion, TMEM9A may play a tumor-promoting role in BC progression via activating the Wnt/β-catenin signaling pathway. Therefore, TMEM9A may be an effective therapeutic option for BC.
INTRODUCTION
Breast cancer (BC) is the most common cancer affecting women and ranks first for incidence in the vast majority of countries.1,2) Current evidence suggests that BC in women aged <45 years is the leading cause of cancer-related deaths.3) According to the report, 20–30% of BC patients may undergo metastasis after tumor resection and adjuvant therapy of the primary tumor.4) Although researchers have made great strides in the treatment and diagnosis of BC,5,6) BC is still a major health issue and currently represents a top biomedical research priority.
Transmembrane proteins (TMEMs) contains an N-terminal signal peptide, a single transmembrane region, three potential N-glycosylation sites, and three conserved cysteine (Cys)-rich domains in the N-terminus.7) TMEM9A, the member of TMEMs family, encodes a 183 amino-acid protein and is highly conserved between species from Caenorhabditis elegans to humans.7) Previous studies indicated that TMEM9A was abnormally expressed in different cancers. For example, TMEM9A was highly expressed in colorectal cancer,8) and it was revealed to be upregulated in regenerating liver and hepatocellular carcinoma cells.9) Furthermore, TMEM9A exerted a tumor-promoting role in human hepatoma cell lines, and TMEM9A knockdown could notably inhibit the proliferation, invasion, migration, and induce apoptosis.10) Another report suggested that TMEM9A could promote intestinal tumorigenesis and hyperactivated the expression of β-catenin.8) Emerging evidence illustrated that TMEM9A was tightly ssociated with cancerous development and exacerbated the malignant behavior of tumors. In this study, bioinformatic data analyzed that TMEM9A expression was upregulated in BC tissues, and we also identified that the transcriptional and protein levels of TMEM9A were prominently increased in BC tissues compared with adjacent normal tissues. However, the function and mechanism of TMEM9A in BC were not well defined.
The Wnt/β-catenin signaling pathway as a conserved signaling pathway is revealed to involve in diverse physiological processes, such as proliferation, differentiation, apoptosis, migration, invasion, and tissue homeostasis.11,12) In the absence of Wnt/β-catenin signal, β-catenin is phosphorylated and hijacked by the destruction complex, which comprises the scaffolding protein Axin, adenomatous polyposis coli (APC), glycogen synthase kinase 3β (GSK-3β), and casein kinase 1α (CK1α).13–16) Subsequently, the phosphorylated β-catenin is recognized and degraded by a ubiquitin ligase-proteasome system.17) Conversely, the Wnt/β-catenin signaling pathway is activated by truncated APC protein, which negates the complex-mediated destruction of β-catenin ubiquitination.18) Once unphosphorylated β-catenin enters the nucleus, it initiates the transcription of downstream genes. Moreover, the Wnt/β-catenin signaling pathway is closely related to the tumorous initiation and progression, which includes cell proliferation and differentiation.19) Thus this pathway exerts important roles in carcinogenesis and therapy response. Khramtsov et al. indicate that wnt/β-catenin pathway activation is enriched in BC and this pathway predicts a lower overall survival rate.20) Therefore, the Wnt/β-catenin pathway is deemed an attractive target for this aggressive BC subtype. In addition, overexpression of TMEM9A enhances the protein expression of the canonical Wnt/β-catenin.21) TMEM9A can activate the Wnt/β-catenin pathway to facilitate liver regeneration and tumorigenesis.9) These findings imply that TMEM9A is closely associated with Wnt/β-catenin signaling, which triggers us to explore whether TMEM9A is involved in the progression of BC by controlling the Wnt/β-catenin pathway.
MATERIALS AND METHODS
Bioinformatic AnalysisThe BC dataset (GSE42568, including 104 breast cancer and 17 normal breast biopsies)22) was downloaded from the Gene Expression Omnibus (GEO, https://www.ncbi.nlm.nih.gov/geoprofiles).23) The Gene Expression Profiling Interactive Analysis (GEPIA) database included BC samples and normal samples was downloaded from GEPIA (http://gepia.cancer-pku.cn).24) In GSE42568, GEO2R tool (http://www.ncbi.nlm.nih.gov/geo/geo2r)25) was used to screen differentially expressed genes (DEGs). For volcano analysis, the standard for screening DEGs was |log2 fold change (FC)| >1.2 and adjusted p < 0.001. Overlapped upregulated genes were screened with log2FC > 1.2 and adjusted p < 0.001 by integrated data, and downregulated genes were screened by the standard of log2FC <−1.2 and adjusted p < 0.001. Overlapped genes were respectively shown in corresponding Venn diagrams (http://bioinformatics.psb.ugent.be/webtools/Venn/). The cluster Profiler R package26) was used to perform the histograms of gene ontology (GO) and Kyoto Encyclopedia of Genes and Genomes (KEGG) in the overlapped genes. The DEGs in TMEM family were screened using the data of GSE42568 and GEPIA. The TMEM9A expression in human BC tissues was analyzed by the TNMplot bioinformatic tool (https://tnmplot.com/analysis/).27) TMEM9A expression in various cancer types was performed by Gene Set Cancer Analysis (http://bioinfo.life.hust.edu.cn/GSCA/#/).28)
BC Tissue SpecimensThirty-seven pairs of para-carcinoma tissues and BC tissues were collected from BC patients as the clinical specimens. Each participant provided informed consent in the study. This study was approved by the Research Ethics Committee of the Fourth Hospital of Hebei Medical University (No: 2020KY113), and this study was carried out following the ethical principles of the latest version of the Declaration of Helsinki.
Cell Culture and TransfectionHS-578T, MDA-MB-231, MDA-MB-468, MCF-7, and MCF-10A cells were purchased from iCell Bioscience (China). T-47D cells were ordered from Procell (China). HS-578T cells and T-47D cells were cultured in Dulbecco's modified Eagle medium and Roswell Park Memorial Institute-1640 medium respectively. Leibovita’s L-15 medium was used to culture MDA-MB-231 and MDA-MB-468 cells. MCF-7 cells were cultured in the minimal essential medium. All of the media were supplemented with 10% fetal bovine serum (FBS) (Tianhang Biotechnology, China) and the above cells were cultured in an incubator at 37 °C and 5% CO2.
T-47D cells were transfected with TMEM9A-specific small interfering RNAs (siRNAs) or the negative control (NC) using Lipofectamine 3000 (Invitrogen, U.S.A.) according to the manufacturer’s protocol. siRNA sequences used in this study were listed in Table 1. The coding sequence (CDS) of TMEM9A was constructed to the plasmid pDONR223 (Youbio Biotechnology, China) to achieve TMEM9A overexpression (TMEM9A-OE). The constructed vector of TMEM9A-OE or empty vector (EV) were transfected into MDA-MB-231 cells using Lipofectamine 3000. For experiments of co-transfection, the reporter plasmids β-Catenin-S33Y (β-Catenin tyrosine for serine at codon 33) or EV and TMEM9A siRNA-4 were co-transfected into T-47D cells, which were named siTMEM9A+β-Catenin-S33Y and siTMEM9A+EV respectively. The transfected cells were used in the follow-up experiments after 48 h incubation.
Table 1. siRNA Sequence Used in the Experiments
| Name | | Sequence (5′–3′) |
|---|
| siTMEM9A-1 | Sense | GCAGCACCACCACCAUCAATT |
| Antisense | UUGAUGGUGGUGGUGCUGCTT |
| siTMEM9A-2 | Sense | GGUGGAAGCUGCAGGUGCATT |
| Antisense | UGCACCUGCAGCUUCCACCTT |
| siTMEM9A-3 | Sense | AGGUCAUCAUUGUCAUCUATT |
| Antisense | UAGAUGACAAUGAUGACCUTT |
| siTMEM9A-4 | Sense | GGCACAAGAUGCUCAGCUATT |
| Antisense | UAGCUGAGCAUCUUGUGCCTT |
| siTMEM9A-5 | Sense | GGAUGCAUAUACUGAGCAATT |
| Antisense | UUGCUCAGUAUAUGCAUCCTT |
| siNC | Sense | ACGUGACACGUUCGGAGAATT |
| Antisense | UUCUCCGAACGUGUCACGUTT |
Total RNA was isolated by TRIpure lysis and reverse-transcribed using BeyoRT II M-MLV reverse transcriptase (Beyotime, China) to synthesize cDNA. For RT-PCR, PCR conditions were 94 °C for 5 min and 35 cycles at 94 °C for 20 s, 55 °C for 20 s and 72 °C for 30 s, followed by a final extension step at 72 °C for 10 min. PCR products were then electrophoresed through a 1.5% agarose gel. The mRNA level of TMEM9A was performed by the gel imaging system. For quantitative RT-PCR, SYBR Green dye (Solarbio, China) was used to measure amplification. The mRNA levels of TMEM9A, cyclin D1, and axis inhibition protein 2 (AXIN2) were analyzed using the 2−ΔΔCT method. Glyceraldehyde-3-phosphate dehydrogenase (GAPDH) was used as endogenous control. The sequences of primers were listed in Table 2.
Table 2. The Sequences of Primers Used in the Experiments
| Name | | Sequence (5′–3′) |
|---|
| TMEM9A | Forward | CTTTGGTGGCTGTGGTC |
| Reverse | CTGGGATACATTCTGGTTG |
| cyclin D1 | Forward | GCGAGGAACAGAAGTGCG |
| Reverse | TGGAGTTGTCGGTGTAGATGC |
| AXIN2 | Forward | ACCGTGGTTGGCTTGTC |
| Reverse | TCTACACTGCTGTCCGTCAT |
| GAPDH | Forward | GACCTGACCTGCCGTCTAG |
| Reverse | AGGAGTGGGTGTCGCTGT |
The whole-cell lysates were collected in radio immunoprecipitation assay (RIPA) lysis buffer (Solarbio, China). The protein concentration was determined using a BCA Protein Assay Kit (Solarbio, China). Equal amounts of protein were separated by electrophoresis and transferred to polyvinylidene difluoride membranes (Millipore, U.S.A.). After blocking with 5% non-fat milk, the membranes were incubated with specific primary antibodies against TMEM9A (Proteintech, China, catalog number: 19918-1-AP, 1 : 1000); APC (Abclnoal, China, catalog number: A17912, 1 : 500); β-catenin (Abclnoal, catalog number: A19657, 1 : 1000); active-β-catenin (CST, China, catalog number: #8814, 1 : 500); GAPDH (Proteintech, catalog number: 60004-1-Ig, 1 : 10000) overnight at 4 °C and subsequently incubated with horseradish peroxidase-conjugated secondary antibody immunoglobulin G (IgG)-horseradish peroxidase (HRP) (Solarbio, catalog number: SE134, 1 : 3000) at 37 °C for 1 h. western blots were detected using ECL Detection Reagents (Solarbio).
3-(4,5-Dimethylthiazol-2-yl)-2,5-diphenyltetrazolium Bromide (MTT)Cells were seeded to a 96-well plate with 7 × 103 cells per well. After 48 h-transfection, the 96-well plates were cultured at 37 °C and 5% CO2 for 0, 24, 48, 72, or 48 h. MTT solution (10 µL, Solarbio) was added to the plate and incubated for 4 h. After removing the supernatant, dissolved in dimethyl sulfoxide (110 µL, Sigma, U.S.A.) was added to dissolve the purple crystals in the dark for 10 min. The optical density was measured on the microplate tester (Biotek, U.S.A.) at 490 nm.
Flow Cytometry AssayFor cell cycle analysis, the transfected cells were fixed with cold 70% ethanol at 4 °C overnight. The cell cycle was detected by a Cell Cycle Analysis Kit (Biosharp, China) according to the manufacturer's protocols. Briefly, the cells were washed with phosphate buffer saline (PBS) and then stained with propidium iodide (PI) at 37 °C for 30 min in the dark. After incubation, the samples were analyzed by a flow cytometer (ACEA Biosciences, U.S.A.).
An apoptosis assay kit (Biosharp) was used to detect cell apoptosis in transfected BC cells. In brief, the cells were washed with PBS and resuspended with 1× Binding buffer. Annexin V-fluorescein isothiocyanate (FITC) (5 µL) was used to stain for 10 min in the dark. After that, PI staining (10 µL) and PBS (400 µL) were added to the cells. The mixed samples were analyzed using flow cytometry.
Wound Healing AssayAfter 48 h-transfection, MDA-MB-231 or T-47D cells were cultured in serum-free medium (SFM) and treated with 1 µg/mL mitomycin C (MCE, U.S.A.) for 1 h. Subsequently, cells were scratched in a straight line with a 200 µL pipette tip and the cell surface was washed with SFM. Each image for the gaps was photographed at the start (0 h) and endpoint (12 h) of the experiment using a microscope (200× magnifications).
TranswellFor invasion assay, transwell was additionally pre-coated with matrigel (Corning, U.S.A.) diluted in a ratio of 1 : 3 with a SFM on ice. The gel was solidified in a 37 °C incubator for 2 h. The transfected cells for the invasion assay were seeded in a SFM in the upper chamber. The lower chamber was supplemented with 10% FBS. After incubation at 37 °C for 24 h, the invasive cells were fixed with paraformaldehyde (Aladdin, China) and stained with 0.1% crystal violet (Amresco, U.S.A.). Finally, cells were photographed and counted using an inverted microscope at 200× magnification (Olympus, Japan).
Statistical AnalysisThe software GraphPad Prism 8.0 was used for statistical analysis. Statistical significance was performed using the one-way ANOVA and the two-tailed Student's t-test. The results were considered significant at p < 0.05 (*), p < 0.01 (**), or p < 0.001 (***). All data were presented as the mean ± standard deviation (S.D.).
RESULTS
Identification of Overlapping DEGs in BC TissuesThe publicly available expression dataset GSE42568 contained the 104 BC and 17 normal breast biopsies were used for bioinformatic analysis. As shown in Fig. 1A, the volcano plot indicated the DEGs in GSE42568 and GEPIA-breast invasive carcinoma (BRCA). The inner track (GSE42568) and outer track (GEPIA) showed the location of the downregulated (green) and upregulated (red) genes on the human chromosome (Fig. 1B). The overlapping upregulated 376 genes and downregulated 587 genes were integrated in GSE42568 and GEPIA, as shown in Venn diagram (Figs. 1C, D). In 376 upregulated genes, GO enrichment analysis were performed, and the terms of top 30 were selected to generate histogram, as shown in Fig. 2A. The KEGG analysis indicated that the upregulated genes were mainly enriched in cell cycle, DNA replication, and bladder cancer (Fig. 2B). The results of GO analysis and KEGG analysis of downregulated genes were shown in Figs. 2C, D. The pathways of KEGG were mainly enriched in peroxisome proliferator-activated receptor (PPAR) signaling pathway, regulation of lipolysis in adipocytes, and pyruvate metabolism.
Identification of Overlapping DEGs in TMEM Family of BC TissuesIt has been reported that TMEM expression is dysregulated and TMEMs as tumor suppressors or oncogenes exhibit oncogenic properties in different tumors.29) Hereon, the volcano plots showed the gene number of TMEM family in GSE42568 and GEPIA (Supplementary Fig. 1). Then the DEGs in TMEM family were screened by overlapping GSE42568 and GEPIA (Fig. 3A). The heat map showed the gene expression in normal and BC tissues of GSE42568, and the results suggested that the number of up-regulated (red) and down-regulated (blue) genes in overlapping TMEM family was 4 and 6, respectively (Fig. 3B). The circos plot showed the 10 genes in the human chromosome. The expression of TMEM9A was analyzed by the data in GSE42568, and it was shown in the center of the circos plot (Fig. 3C). By analyzing the expression of TMEM9A in BC, we found that TMEM9A was significantly upregulated in tumor tissues compared with tumor-adjacent samples (Fig. 3D) and normal samples (Fig. 3E). Figure 3F showed the dysregulated TMEM9A expression in different cancer types, and we discovered that TMEM9A was upregulated in BRCA samples.
The Expression of TMEM9A Is Upregulated in BC Tissues and CellsThe increased mRNA level of TMEM9A was validated using qRT-PCR in 37 pairs of para-carcinoma tissues and BC tissues (Fig. 4A). Subsequently, 6 pairs of samples were used for the detection of the transcriptional and protein levels of TMEM9A (Fig. 4B). The results indicated that TMEM9A was profoundly upregulated in BC tissues compared with para-carcinoma tissues. Normal cell line (MCF-10A cells), and BC cell lines (HS-578T, MDA-MB-231, MDA-MB-468, MCF-7, and T-47D cells) were used to identify the mRNA and protein levels of TMEM9A. The results of RT-PCR and Western blot showed that TMEM9A was highly expressed in BC cell lines compared with the MCF-10A line (Figs. 4C, D). To study the biological function of TMEM9A in BC, we transfected TMEM9A-OE and si-TMEM9A to MDA-MB-231 (the lowest TMEM9A expression) and T-47D cells (the highest TMEM9A expression) respectively. The results of qRT-PCR and Western blot verified the transfected efficiency in MDA-MB-231 cells and T-47D cells (Figs. 4E–H). Therein, si-TMEM9A-4 and si-TMEM9A-5 were selected for the followed experiments (Figs. 4F, H).
Modulation of TMEM9A Expression Alters Cell Cycle and Apoptosis in BC CellsAs indicated in Figs. 5A and B, the MTT assay suggested that TMEM9A knockdown notably decreased the cell viability compared with the NC group at 24 h, 48 h, and 96 h, whereas TMEM9A overexpression exerted the opposite effects. The percentage of cells in the G1 phase in the TMEM9A-OE group was markedly decreased, and the cell distribution in the G2 and S phases were increased (Fig. 5C). The flow cytometric analysis revealed that TMEM9A knockdown lead to cell cycle arrest in T-47 cells (Fig. 5D, Supplementary Fig. 2). Also, the apoptotic rate was higher in TMEM9A-silenced cells than in control cells (Fig. 5E).
Modulation of TMEM9A Expression Promotes Migration and Invasion in BC CellsTo further probe the impact of TMEM9A on malignant behaviors, we detected the migratory and invasive abilities in BC cells. Wound healing assay showed that TMEM9A overexpression promoted migration, and TMEM9A deficiency prominently inhibited the migratory activity (Figs. 6A, C). The results of the transwell assay suggested that the overexpression of TMEM9A markedly enhanced cell invasion in MDA-MB-231 cells, contrarily, the invasive ability was decreased in the TMEM9A-silenced cells (Figs. 6B, D).
TMEM9A Affects the Activation of the Wnt/β-Catenin Signaling PathwayGiven that the association between the Wnt/β-catenin signaling and carcinoma as well as TMEM9A, we explored the effects of TMEM9A on the Wnt/β-catenin pathway in BC cells. Western blot analysis suggested that TMEM9A overexpression presented the upregulated expression of total and active β-catenin, while the protein level of APC was dropped, and TMEM9A deficiency exerted the opposite effects (Figs. 7A, B). It could be seen from Figs. 7C, E that the mRNA level of cyclin D1 and AXIN2 was increased in TMEM9A-overexpressed MDA-MB-231 cells, whereas TMEM9A deletion decreased the expression of cyclin D1 and AXIN2 in T-47D cells (Figs. 7D, F).
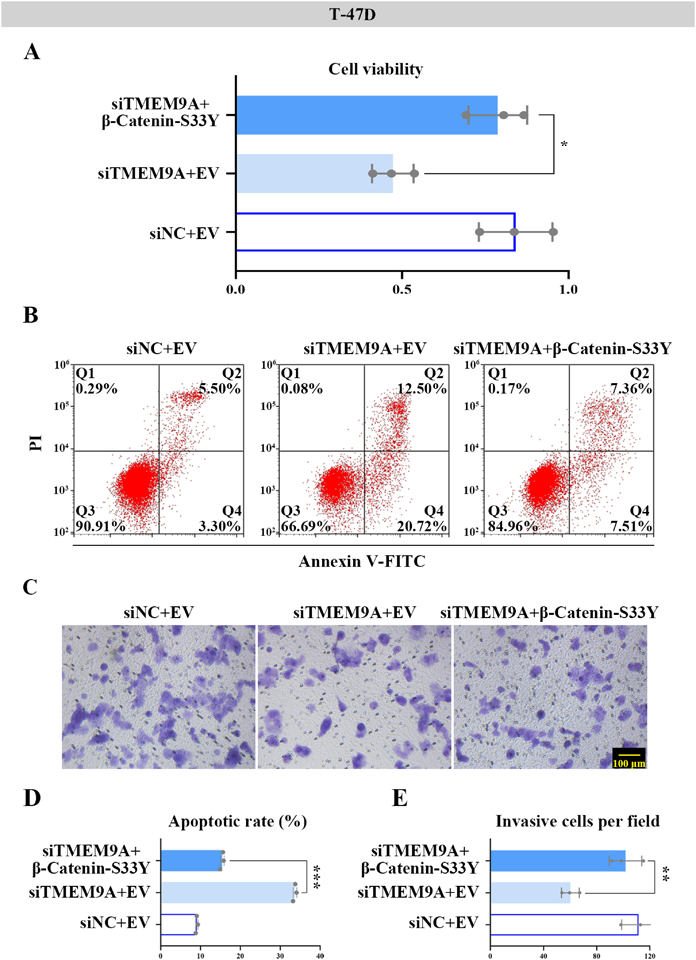
TMEM9A Regulates the Development of BC Cells through the Wnt/β-Catenin PathwayPhosphorylated β-catenin is degraded after it is dissociated from the destruction complex, while un-phosphorylated β-catenin can migrate to the nucleus and accumulate, thereby triggering the Wnt/β-catenin pathway.19,30) T-47D cells with TMEM9A deletion were transfected with the plasmid of β-Catenin-S33Y (non-phosphorylated status) to activate the Wnt/β-catenin pathway. The results of MTT assay suggested that the siTMEM9A + EV group decreased the cell viability, which was reversed by the siTMEM9A + β-Catenin-S33Y group (Fig. 8A). The elevated apoptotic rate in T-47D cells induced by TMEM9A knockdown was suppressed in the presence of β-Catenin-S33Y (Figs. 8B, D). As shown in Figs. 8C and E, the decreased invasive cells after TMEM9A silencing were partially rescued via β-catenin activation.
DISCUSSION
TMEMs are reported to be upregulated or downregulated in cancerous tissues compared to adjacent normal tissues.29) TMEMs also participate in drug resistance, and some of TMEMs are used as prognostic biomarkers,29,31) suggesting that TMEM family as a prominent group plays the important role in cancer research. In this study, bioinformatics analysis identified the overlapped DEGs in TMEMs and their expression was shown in the heat map. Hereon, upregulated TMEM family genes were been selected to investigate the tumor-promoting role in BC. Among the four up-regulated genes, TMEM97 is reported to involve in the proliferation and migration of the BC cell lines.32,33) According to the published literatures and the basis of the thesis, we preferentially selected TMEM9A as a candidate gene, and we will further explore the function of the other members of TMEM family in BC in future studies. Studies have demonstrated that the expression of TMEM9A was associated with the function of a series of cancers such as hepatocellular carcinoma and colorectal cancer.8,10) This study summarized the findings and contributions of TMEM9A to BC and demonstrated that TMEM9A was highly expressed in BC tissues and cell lines. Considering that TMEM9A may be a putative oncogene, silencing or expressing its expression would result in therapeutic effects. In this study, the significant expression of TMEM9A confirmed the transfected efficiency, thus, we did not conduct experiments to verify the percentage of cell transfection efficiency. In vitro experiments showed that genetic ablation of TMEM9A inhibited cell phenotypes, whereas TMEM9A overexpression presented the opposite effects. A strong relationship between TMEM9A and malignant behavior has been described in hepatocellular carcinoma.10)
Wnt/β-catenin signaling involves multiple cellular processes, including proliferation, metastasis, immune microenvironment regulation, and phenotype shaping of BC.34–37) Evidence increasingly has demonstrated that the transcription factor β-catenin is a key component of the Wnt/β-catenin signaling pathway and abnormal regulation of β-catenin can lead to the occurrence of early cancer events.19,38,39) Russell et al. have manifested that cyclin D1, the targeting protein of the β-catenin pathway, may promote hepatocyte proliferation.40) Our studies suggested that TMEM9A predominantly affected the mRNA levels of cyclin D1 and AXIN2, the target genes in the downstream Wnt/β-catenin signaling pathway. Additionally, TMEM9A downregulated the protein production of APC and enriched the protein amounts of β-catenin and active β-catenin, uncovering that TMEM9A accelerated the activation of the Wnt/β-catenin pathway. Based on the findings, we characterized the role of the TMEM9A-activated Wnt/β-catenin pathway in BC development. It is well known that β-Catenin-S33Y is insensitive to GSK-3β-mediated phosphorylation and proteasomal degradation.41) Then we found that β-Catenin-S33Y rescued the decreased cell viability and invasion caused by TMEM9A knockdown in BC cells. Loss of TMEM9A increased the apoptotic rate, while the stimulation of β-catenin reversed the status, revealing that TMEM9A could control the Wnt/β-catenin pathway to promote BC development. Researchers have demonstrated that TMEM9A affects liver regeneration by downregulating APC through lysosomal protein degradation, and further hyperactivating the Wnt/β-catenin pathway,9) which is consistent with the present results. Our study also supports evidence from the exploration reported by Jung et al. that TMEM9A facilitates intestinal tumorigenesis by Wnt/β-catenin signaling pathway.8)
Several reviews have comprehensively discussed the important role of the Wnt/β-catenin pathway in tumorous development and therapy.19,42,43) It is well known that inflammation is recognized as a hallmark feature of cancer progression.44) In the last few decades, the impact of inflammation and the immune system on cancerous progression has gained extensive attention. In inflammatory cancers, the Wnt/β-catenin signaling pathway has emerged as a key pathway in malignant biology.45,46) Suryawanshi et al. report the effect on the mutual coexistence and co-dependence of the Wnt/β-catenin signaling pathway along with various inflammatory cascades.47) A novel strategy for cancer prevention and therapy targeting the crosstalk between canonical Wnt/β-catenin and inflammatory signaling cascades has been mentioned.48) The current study indicated that TMEM9A regulated the Wnt/β-catenin pathway to accelerate breast neoplastic progression. Given the connection between inflammation and cancer as well as the Wnt/β-catenin pathway, we have a conjecture that TMEM9A might also affect the occurrence of inflammation. In hepatoma cells, TMEM9A has been demonstrated to mediate interleukin (IL)-6 and IL-1β secretion as well as modulate the Wnt/β-catenin signaling pathway.21) The evidence suggests that the inflammatory components are involved in neoplastic processes, including proliferation, survival, invasion, and migration, making inflammation serves the primary target for cancer prevention and treatment.48) As a novel human transmembrane protein, TMEM9A and its function referred to the BC-related inflammatory response were needed to further explore. There is abundant room for further progress in determining the role of TMEM9A in BC progression.
In summary, this study provided evidence for the first time that TMEM9A was significantly expressed in BC samples, and manifested that TMEM9A played an important role in the cell proliferation, migration, and apoptosis of BC cells. Overall, by regulating the Wnt/β-catenin signaling pathway, TMEM9A exerts the tumor-promoting properties on the development of BC cells, which may be useful for the therapy of BC.
Conflict of Interest
The authors declare no conflict of interest.
Supplementary Materials
This article contains supplementary materials.
REFERENCES
- 1) Ferlay J, Soerjomataram I, Dikshit R, Eser S, Mathers C, Rebelo M, Parkin DM, Forman D, Bray F. Cancer incidence and mortality worldwide: sources, methods and major patterns in GLOBOCAN 2012. Int. J. Cancer, 136, E359–E386 (2015).
- 2) Sung H, Ferlay J, Siegel RL, Laversanne M, Soerjomataram I, Jemal A, Bray F. Global cancer statistics 2020: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J. Clin., 71, 209–249 (2021).
- 3) Anastasiadi Z, Lianos GD, Ignatiadou E, Harissis HV, Mitsis M. Breast cancer in young women: an overview. Updates Surg., 69, 313–317 (2017).
- 4) Chaffer CL, Weinberg RA. A perspective on cancer cell metastasis. Science, 331, 1559–1564 (2011).
- 5) Appierto V, Di Cosimo S, Reduzzi C, Pala V, Cappelletti V, Daidone MG. How to study and overcome tumor heterogeneity with circulating biomarkers: the breast cancer case. Semin. Cancer Biol., 44, 106–116 (2017).
- 6) Rossi T, Gallerani G, Martinelli G, Maltoni R, Fabbri F. Circulating tumor cells as a tool to untangle the breast cancer heterogeneity issue. Biomedicines, 9, 1242 (2021).
- 7) Kveine M, Tenstad E, Døsen G, Funderud S, Rian E. Characterization of the novel human transmembrane protein 9 (TMEM9) that localizes to lysosomes and late endosomes. Biochem. Biophys. Res. Commun., 297, 912–917 (2002).
- 8) Jung Y-S, Jun S, Kim MJ, Lee SH, Suh HN, Lien EM, Jung H-Y, Lee S, Zhang J, Yang J-I, Ji H, Wu JY, Wang W, Miller RK, Chen J, McCrea PD, Kopetz S, Park J-I. TMEM9 promotes intestinal tumorigenesis through vacuolar-ATPase-activated Wnt/β-catenin signalling. Nat. Cell Biol., 20, 1421–1433 (2018).
- 9) Jung Y-S, Stratton SA, Lee SH, Kim M-J, Jun S, Zhang J, Zheng B, Cervantes CL, Cha J-H, Barton MC, Park J-I. TMEM9-v-ATPase activates Wnt/β-catenin signaling via APC lysosomal degradation for liver regeneration and tumorigenesis. Hepatology, 73, 776–794 (2021).
- 10) Zhang Y, Ran Y, Xiong Y, Zhong Z-B, Wang Z-H, Fan X-L, Ye Q-F. Effects of TMEM9 gene on cell progression in hepatocellular carcinoma by RNA interference. Oncol. Rep., 36, 299–305 (2016).
- 11) Choi B-R, Cave C, Na CH, Sockanathan S. GDE2-dependent activation of canonical Wnt signaling in neurons regulates oligodendrocyte maturation. Cell Reports, 31, 107540 (2020).
- 12) Soleas JP, D’Arcangelo E, Huang L, Karoubi G, Nostro MC, McGuigan AP, Waddell TK. Assembly of lung progenitors into developmentally-inspired geometry drives differentiation via cellular tension. Biomaterials, 254, 120128 (2020).
- 13) Rubinfeld B, Albert I, Porfiri E, Fiol C, Munemitsu S, Polakis P. Binding of GSK3beta to the APC-beta-catenin complex and regulation of complex assembly. Science, 272, 1023–1026 (1996).
- 14) Hart MJ, de los Santos R, Albert IN, Rubinfeld B, Polakis P. Downregulation of beta-catenin by human Axin and its association with the APC tumor suppressor, beta-catenin and GSK3 beta. Curr. Biol., 8, 573–581 (1998).
- 15) Yost C, Torres M, Miller JR, Huang E, Kimelman D, Moon RT. The axis-inducing activity, stability, and subcellular distribution of beta-catenin is regulated in Xenopus embryos by glycogen synthase kinase 3. Genes Dev., 10, 1443–1454 (1996).
- 16) Amit S, Hatzubai A, Birman Y, Andersen JS, Ben-Shushan E, Mann M, Ben-Neriah Y, Alkalay I. Axin-mediated CKI phosphorylation of beta-catenin at Ser 45: a molecular switch for the Wnt pathway. Genes Dev., 16, 1066–1076 (2002).
- 17) Aberle H, Bauer A, Stappert J, Kispert A, Kemler R. beta-catenin is a target for the ubiquitin–proteasome pathway. EMBO J., 16, 3797–3804 (1997).
- 18) Vermeulen L, De Sousa E Melo F, van der Heijden M, Cameron K, de Jong JH, Borovski T, Tuynman JB, Todaro M, Merz C, Rodermond H, Sprick MR, Kemper K, Richel DJ, Stassi G, Medema JP. Wnt activity defines colon cancer stem cells and is regulated by the microenvironment. Nat. Cell Biol., 12, 468–476 (2010).
- 19) Zhang X, Wang L, Qu Y. Targeting the β-catenin signaling for cancer therapy. Pharmacol. Res., 160, 104794 (2020).
- 20) Khramtsov AI, Khramtsova GF, Tretiakova M, Huo D, Olopade OI, Goss KH. Wnt/beta-catenin pathway activation is enriched in basal-like breast cancers and predicts poor outcome. Am. J. Pathol., 176, 2911–2920 (2010).
- 21) Wei W, Jiang F, Liu X-C, Su Q. TMEM9 mediates IL-6 and IL-1β secretion and is modulated by the Wnt pathway. Int. Immunopharmacol., 63, 253–260 (2018).
- 22) Clarke C, Madden SF, Doolan P, Aherne ST, Joyce H, O’Driscoll L, Gallagher WM, Hennessy BT, Moriarty M, Crown J, Kennedy S, Clynes M. Correlating transcriptional networks to breast cancer survival: a large-scale coexpression analysis. Carcinogenesis, 34, 2300–2308 (2013).
- 23) Edgar R, Domrachev M, Lash AE. Gene Expression Omnibus: NCBI gene expression and hybridization array data repository. Nucleic Acids Res., 30, 207–210 (2002).
- 24) Tang Z, Li C, Kang B, Gao G, Li C, Zhang Z. GEPIA: a web server for cancer and normal gene expression profiling and interactive analyses. Nucleic Acids Res., 45 (W1), W98–W102 (2017).
- 25) Barrett T, Wilhite SE, Ledoux P, Evangelista C, Kim IF, Tomashevsky M, Marshall KA, Phillippy KH, Sherman PM, Holko M, Yefanov A, Lee H, Zhang N, Robertson CL, Serova N, Davis S, Soboleva A. NCBI GEO: archive for functional genomics data sets--update. Nucleic Acids Res., 41 (D1), D991–D995 (2013).
- 26) Yu G, Wang L-G, Han Y, He Q-Y. clusterProfiler: an R package for comparing biological themes among gene clusters. OMICS, 16, 284–287 (2012).
- 27) Bartha Á, Győrffy B. TNMplot.com: A web tool for the comparison of gene expression in normal, tumor and metastatic tissues. Int. J. Mol. Sci., 22, 2622 (2021).
- 28) Liu C-J, Hu F-F, Xia M-X, Han L, Zhang Q, Guo A-Y. GSCALite: a web server for gene set cancer analysis. Bioinformatics, 34, 3771–3772 (2018).
- 29) Schmit K, Michiels C. TMEM proteins in cancer: a review. Front Pharmacol, 9, 1345 (2018).
- 30) Stamos JL, Weis WI. The β-catenin destruction complex. Cold Spring Harb. Perspect. Biol., 5, a007898 (2013).
- 31) Wrzesiński T, Szelag M, Cieślikowski WA, Ida A, Giles R, Zodro E, Szumska J, Poźniak J, Kwias Z, Bluyssen HAR, Wesoly J. Expression of pre-selected TMEMs with predicted ER localization as potential classifiers of ccRCC tumors. BMC Cancer, 15, 518 (2015).
- 32) Cantonero C, Camello PJ, Salido GM, Rosado JA, Redondo PC. TMEM97 facilitates the activation of SOCE by downregulating the association of cholesterol to Orai1 in MDA-MB-231 cells. Biochim. Biophys. Acta BBAMol. Cell Biol. Lipids, 1866, 158906 (2021).
- 33) Cantonero C, Camello PJ, Abate C, Berardi F, Salido GM, Rosado JA, Redondo PC. NO1, a new sigma 2 receptor/TMEM97 fluorescent ligand, downregulates SOCE and promotes apoptosis in the triple negative breast cancer cell lines. Cancers (Basel), 12, 257 (2020).
- 34) Wend P, Runke S, Wend K, Anchondo B, Yesayan M, Jardon M, Hardie N, Loddenkemper C, Ulasov I, Lesniak MS, Wolsky R, Bentolila LA, Grant SG, Elashoff D, Lehr S, Latimer JJ, Bose S, Sattar H, Krum SA, Miranda-Carboni GA. WNT10B/β-catenin signalling induces HMGA2 and proliferation in metastatic triple-negative breast cancer. EMBO Mol. Med., 5, 264–279 (2013).
- 35) Luga V, Zhang L, Viloria-Petit AM, Ogunjimi AA, Inanlou MR, Chiu E, Buchanan M, Hosein AN, Basik M, Wrana JL. Exosomes mediate stromal mobilization of autocrine Wnt-PCP signaling in breast cancer cell migration. Cell, 151, 1542–1556 (2012).
- 36) Malladi S, Macalinao DG, Jin X, He L, Basnet H, Zou Y, de Stanchina E, Massagué J. Metastatic latency and immune evasion through autocrine inhibition of WNT. Cell, 165, 45–60 (2016).
- 37) Jiang S, Zhang M, Zhang Y, Zhou W, Zhu T, Ruan Q, Chen H, Fang J, Zhou F, Sun J, Yang X. WNT5B governs the phenotype of basal-like breast cancer by activating WNT signaling. Cell Commun. Signal., 17, 109 (2019).
- 38) Lim Z-F, Ma PC. Emerging insights of tumor heterogeneity and drug resistance mechanisms in lung cancer targeted therapy. J. Hematol. Oncol., 12, 134 (2019).
- 39) Zhou J, Toh SH-M, Chan Z-L, Quah JY, Chooi J-Y, Tan TZ, Chong PSY, Zeng Q, Chng W-J. A loss-of-function genetic screening reveals synergistic targeting of AKT/mTOR and WTN/β-catenin pathways for treatment of AML with high PRL-3 phosphatase. J. Hematol. Oncol., 11, 36 (2018).
- 40) Russell JO, Monga SP. Wnt/β-catenin signaling in liver development, homeostasis, and pathobiology. Annu. Rev. Pathol., 13, 351–378 (2018).
- 41) Kolligs FT, Hu G, Dang CV, Fearon ER. Neoplastic transformation of RK3E by mutant beta-catenin requires deregulation of Tcf/Lef transcription but not activation of c-myc expression. Mol. Cell. Biol., 19, 5696–5706 (1999).
- 42) Zhang Y, Wang X. Targeting the Wnt/β-catenin signaling pathway in cancer. J. Hematol. Oncol., 13, 165 (2020).
- 43) Krishnamurthy N, Kurzrock R. Targeting the Wnt/beta-catenin pathway in cancer: update on effectors and inhibitors. Cancer Treat. Rev., 62, 50–60 (2018).
- 44) Hanahan D, Weinberg RA. Hallmarks of cancer: the next generation. Cell, 144, 646–674 (2011).
- 45) An X, Liu X, Zhang L, Liu J, Zhao X, Chen K, Ma H, Li G, Cao B, Song Y. MiR-449a regulates caprine endometrial stromal cell apoptosis and endometrial receptivity. Sci. Rep., 7, 12248 (2017).
- 46) Polakis P. Wnt signaling in cancer. Cold Spring Harb. Perspect. Biol., 4, a008052 (2012).
- 47) Suryawanshi A, Tadagavadi RK, Swafford D, Manicassamy S. Modulation of inflammatory responses by Wnt/β-catenin signaling in dendritic cells: a novel immunotherapy target for autoimmunity and cancer. Front. Immunol., 7, 460 (2016).
- 48) Tewari D, Bawari S, Sharma S, DeLiberto LK, Bishayee A. Targeting the crosstalk between canonical Wnt/β-catenin and inflammatory signaling cascades: a novel strategy for cancer prevention and therapy. Pharmacol. Ther., 227, 107876 (2021).