Technical paper
Improved Cultivation and Fortification of The Giant Embryo Brown Rice Cultivar “Koshiguruma”
2018 Volume 24 Issue 4 Pages 619-626
Details
2018 Volume 24 Issue 4 Pages 619-626
In general, giant embryo rice has a lower germination rate and shows non-uniformity in seedling emergence. We attempted to determine suitable pH and temperature conditions for germinating the giant embryo rice cultivar Koshiguruma, grown in Niigata Prefecture, in combination with red onion extract. We also examined metabolic changes in the grains that could have important human health benefits. Koshiguruma showed a higher germination rate (1.1 times) when germinated with red onion extract for 24 h at 37 °C and contained more gamma aminobutyric acid (1.7 times) than when germinated with water. In addition, germination of Koshiguruma in combination with polyphenols imparted antioxidant capacities. Thus, we have established methods for improved cultivation of giant embryo rice, which may have important human health benefits upon consumption.
Rice is a primary staple food in Asia. However, rice consumption has declined in Japan since 1974 due to changes in dietary habits. The annual per capita rice consumption was 111.7 kg in 1965, declining to 54.4 kg in 2016, which is a reduction of more than 50 % (Ministry of Agriculture, Forestry and Fisheries, 2017). As a consequence, the Ministry of Agriculture, Forestry and Fisheries instituted the Super Rice Project in 1989 to develop rice cultivars with new characteristics, for example, low- or high-amylose rice cultivars, red or purple pigmented rice, and giant embryo rice (Kobayashi, 1994).
Although it is known that brown rice grains contain greater nutritional benefits than ordinary milled rice grains, including increased dietary fiber, phytic acid, vitamins B and E, and gamma aminobutyric acid (GABA), brown rice is not considered suitable as table rice because of its dark appearance and hard texture. The texture of pre-germinated brown rice can be improved by the activation of enzymes that digest the starch and cell walls. As a consequence, the Glycemic Index (GI) value of pre-germinated brown rice is about 60 % that of ordinary white rice (Ohtsubo et al., 2005).
By crossing Hokkai 269 gou and Niigatawase cultivars, the Niigata Prefectural Agricultural Research Institute developed a non-glutinous cultivar in 2008 that contains a giant embryo, known as Koshiguruma (Kobayashi et al., 2009). Koshiguruma is a very short-culmed variety, with short ears, and is not heavy (1000-kernel weight). In ordinary rice cultivars, the embryo weight occupies about 2 % of the whole grain weight; however, the embryo volume of Koshiguruma is 2.5–2.8 times that of ordinary rice cultivars. In addition, pre-germinated brown Koshiguruma rice has been reported to have important health benefits, such as reducing blood pressure, postprandial blood glucose, and insulin concentration (Watanabe et al., 2015). However, it is characterized by late seedling emergence and non-uniform germination compared to Koshihikari, leading to poor seedling establishment during transplantation and cultivation of Koshiguruma (Kobayashi et al., 2004). In addition, pre-germinated brown rice is rich in nutritional components, leading to rampant infestation by microorganisms.
Koshiguruma has limitations with respect to germination, but pre-germinated brown rice has been reported to show beneficial effects as described before. Thus, pre-germinated Koshiguruma consumption may be beneficial in preventing hypertension and diabetes mellitus.
Saikusa et al. (1994) reported that GABA was increased during soaking of rice germs and brown rice grains via alterations in glutamic acid (Saikusa et al., 1994a; Saikusa et al., 1994b). GABA is a non-protein amino acid that is widely distributed in nature and is known to exist in the brains of vertebrates as a suppressive neurotransmitter (Awapara et al., 1950; Roberts, 1950). It is effective for alleviating increases in blood pressure (Omori et al., 1987; Shirai et al., 2011; Yasui et al., 2004), mental symptoms in menopausal and presenile autonomic nervous disorders (Okuda et al., 2000), stress (Mori et al., 2007), growth hormone secretion (Cavagnini et al., 1980), kidney function disorder (Hyun et al., 2004; Lin et al., 2000), neutral fat increase (Ohtomo, 2010), and more.
To address the problems of poor germination in pre-germinated brown rice, including poor germination rate, seedling establishment, and microorganism infestation, Nakamura and Ohtsubo (2010) reported that red onion extract had germination-promoting and antibacterial effects. Red onion, which we used in the present investigation, contains high levels of quercetin and its derivatives (Nakamura and Ohtsubo, 2010). Quercetin is the aglycone of numerous flavonoid glycosides, such as rutin and quercitrin, and is commonly found in citrus fruit, buckwheat and onions. Quercetin has been reported to have anti-inflammatory, antimutagenic and antioxidant properties (Kobori et al., 1999; Singh et al., 2009).
Therefore, we examined the effects of red onion extract on the pre-germinated giant embryo rice cultivar Koshiguruma. In particular, we investigated whether red onion extract served as a suitable soaking solution to improve germination conditions, as well as to increase GABA, polyphenols and anti-oxidant contents. Furthermore, we examined endogenous enzyme activities during germination to assess the effects of red onion extract.
Rice samples We used three rice cultivars with different nutritional characteristics. “Koshiguruma” is a giant embryo rice cultivar, “Koshihikari” is a high-quality premium rice cultivar, and “Hoshiyutaka” is a high-amylose rice cultivar. Koshiguruma and Koshihikari were provided by Niigata Crop Research Center (harvested in 2010), and Hoshiyutaka was provided by Kyushu University (harvested in 2010). The brown rice was polished to 90 % (w/w) using a grinding rice mill (Test Mill TM05C; Satake Co., Ltd., Higashihiroshima, Japan). White rice flour was prepared using a cyclone mill (SFC-SI; Udy, Fort Collins, CO, USA) with a screen containing 1 mm diameter pores.
Suitable pH and temperature of solution for germination To determine the suitable solution temperature for germination, 3-g portions of brown rice samples were soaked in water and incubated at 30, 35, and 40 °C for 18 h. To determine the suitable pH of the solution for germination, 3-g portions of brown rice samples were soaked in water for 18 h at pH 4.0, 5.0, 6.0, or 7.0. After incubation, the germination rate was calculated. Each treatment was performed in triplicate.
Preparation of water and red onion solution for germination Water and red onion solution for soaking were prepared using the germination conditions obtained from the preliminary experiment (Koshiguruma: pH 5.5, 37 °C; Koshihikari: pH 5.6, 36 °C; Hoshiyutaka: pH 6.5, 35 °C). Based on the report by Nakamura and Ohtsubo (2010), the concentration of the red onion solution was adjusted to 2 %.
Preparation of germinated rice samples To investigate the effect of germination, we prepared samples germinated for 6, 18, 24, 30, 36, 48, 60, and 72 h. Each rice cultivar was soaked in water or red onion solution. The germination rate was calculated, and the geminated brown rice samples were lyophilized and pulverized using a coffee mill (IMF-650D; Iwatani, Tokyo, Japan) to measure the GABA content, etc.
Analyses of general components The moisture content was determined by the oven-dry method (135 °C, 1 h), and the amylose content was determined using the iodine colorimetric method developed by Juliano (1971).
Measurement of GABA content The GABA content of rice flour was determined using an amino acid analyzer (Amino Tac JLC-500/v; JEOL Ltd., Tokyo, Japan). To measure GABA, 0.6 g of rice flour was extracted for 20 min by shaking in 5 mL of a 10 % sulfosalicylic acid solution at room temperature. The extract was adjusted to pH 2.2 with 3 M NaOH, and then the solution was adjusted to 10 mL with sodium citrate buffer, followed by centrifugation at 2000 rpm for 10 min. Finally, the supernatant was filtered through a 0.45 µm membrane filter and the GABA content was measured using an amino acid analyzer.
Measurement of polyphenol content The polyphenol content of rice flour was determined using the Folin-Ciocalteu method (Tsuji and Kimura, 2001). To measure the polyphenol content, 200 mg of rice flour was extracted for 30 min with 4 mL of 50 % EtOH at room temperature, after which the extract was centrifuged at 3500 rpm for 15 min. Equal volumes of water and Folin-Ciocalteu solution were added to 0.5 mL of the supernatant, mixed, and then incubated at room temperature for 3 min. Subsequently, 5 mL of sodium carbonate was added, and the solution was incubated at 50 °C for 5 min. The sample solution was cooled and allowed to stand for 1 h at 10 °C. The absorbance was measured at 765 nm, and gallic acid (0.1 mg/mL) was used for calibration.
Measurement of DPPH radical scavenging capacity The antioxidative activity of rice flour was determined using the method described by Kimura et al. (2008). To measure the DPPH radical scavenging capacity, 200 mg of rice flour was extracted for 30 min by shaking with 4 mL of ethanol at room temperature and then centrifuged at 3000 rpm for 10 min. The supernatant (0.5 mL) and 0.5 mL of buffer A [0.4 mM DPPH (2,2-diphenyl 1-1-picrylhydrazyl: 0.2 M MES (pH 6.0): 20 % ethanol=, mixed at 1:1:1) were mixed at room temperature for 20 min in a dark room. Another 0.5 mL of supernatant was mixed with 0.5 mL of buffer B [99.5 % EtOH: 0.2 M MES (pH 6.0): 20 % ethanol=, mixed at 1:1:1] at room temperature for 20 min in a dark room. The absorbance was measured at 520 nm. Trolox (0.042 mM) was used for calibration. The absorbance corresponding to the DPPH radical scavenging capacity was calculated as the difference between A (the absorbance in the case of buffer A) and B (the absorbance in the case of buffer B).
α-Amylase activity The α-amylase activity of rice flour samples was determined using an enzyme assay kit (Megazyme, Wicklow, Ireland). To measure the α-amylase activity, 10 mg of rice flour was extracted for 30 min with 1 mL of extraction buffer at pH 5.4 and room temperature, after which the extract was centrifuged at 2930 rpm for 10 min. The extract solution (0.1 mL) and substrate (0.1 mL) were pre-incubated at 40 °C for 5 min. Each solution was then incubated at 40 °C for exactly 20 min before the addition of stopping reagent (1.5 mL). The absorbance was measured at 400 nm.
Cellulase activity The cellulase activity of the milled rice flour samples was determined using an enzyme assay kit (Megazyme). To measure cellulase activity, 375 mg of rice flour was extracted for 15 min at room temperature with 6 mL of 100 mM sodium acetate buffer at pH 4.5, after which the extract was centrifuged at 2930 rpm for 10 min. The extract solution (0.5 mL) and Azo-Barley glucan substrate solution (0.5 mL) were pre-incubated at room temperature for 5 min. Each solution was incubated at room temperature for exactly 10 min before the addition of stopping reagent (3 mL). The absorbance was measured at 590 nm.
Statistical analysis All the results, including the significance of regression coefficients, were subjected to statistical analyses using t-tests and one-way ANOVA according to Tukey's method, and were performed in Excel Statistics (ver. 2006; Microsoft Corporation, Tokyo, Japan).
Moisture and amylose contents of rice samples The moisture content of brown rice and amylose content of milled rice are shown in Table 1. Koshiguruma showed a slightly higher amylose content than Koshihikari, but Hoshiyutaka showed the highest content, consistent with a previous report (Shinoda et al., 1990).
| Moisture content (%) | Amylose content (%) | |
|---|---|---|
| Koshiguruma | 12.2±0.02 | 19.6±0.46 |
| Koshihikari | 12.3±0.28 | 16.5± 0.46 |
| Hoshiyutaka | 11.3±0.23 | 30.1±0.46 |
Suitable pH and temperature of soaking water for germination The suitable temperature and pH for soaking the rice cultivars were determined. All seeds showed a high germination rate at 35 °C and pH 5.0 to 6.0 (Fig. 1). Therefore, an estimation formula was prepared, and the temperature and pH at which the germination rate became the highest were calculated and used in the following experiments.

Effects of pH and temperature on germination rate. The diamond denotes Koshiguruma, the square refers to Hoshiyutaka, and triangle indicates Koshihikari. The line connecting the points shows the approximate curve.
Change in germination rate Almost all grains germinated after soaking for 24 h, while Koshiguruma and Koshihikari showed the best germination after soaking for 36 h (Fig. 2). Although the germination rate of giant embryo rice was reported to be slow (Uehara et al., 2003), Koshiguruma showed an equivalent germination rate to Koshihikari, and the germination rate increased further by soaking in red onion solution. This result was consistent with that reported by Nakamura and Ohtsubo (2010). To elucidate the accelerating action of red onion extract on germination, uniconazole, which is a synthetic inhibitor of gibberellic acid (GA), was added to the extract and samples were germinated. Samples treated with red onion solution containing uniconazole (data not shown) showed reduced germination success, suggesting that acceleration of germination with red onion extract is likely caused by an auxin-like substance. Pradhan and Basu reported that quercetin promoted germination of leguminous plant seeds as a cofactor of indole acetic acid (IAA) at extremely low concentrations (Pradhan and Basu, 1981). In this context, the authors considered that the germination promoting action of quercetin is cultivar specific, and that Koshiguruma showed accelerated germination by the quercetin contained in red onion.
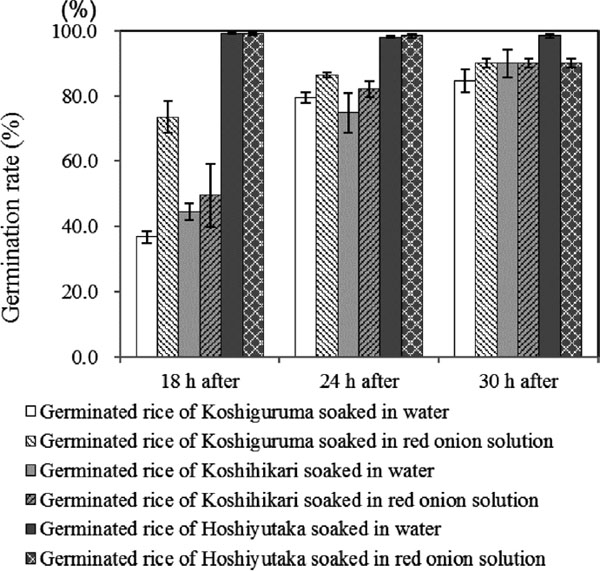
Germination rate of each rice samples soaked either in red onion solution or water.  refers to germinated rice of Koshiguruma soaked in water,
refers to germinated rice of Koshiguruma soaked in water,  shows germinated rice of Koshiguruma soaked in red onion solution.
shows germinated rice of Koshiguruma soaked in red onion solution.  shows germinated rice of Koshihikari soaked in water,
shows germinated rice of Koshihikari soaked in water,  shows germinated rice of Koshihikari soaked in red onion solution,
shows germinated rice of Koshihikari soaked in red onion solution,  shows germinated rice of Hoshiyutaka soaked in water, and
shows germinated rice of Hoshiyutaka soaked in water, and  shows germinated rice of Hoshiyutaka soaked in red onion solution.
shows germinated rice of Hoshiyutaka soaked in red onion solution.
GABA content The GABA content of the germinated rice samples is shown in Table 2. There are many reports on the enhancing effects of increased GABA on the germination rate (Izumi et al., 1984; Nakamura and Ohtsubo, 2010; Phantipha et al., 2009; Saikusa et al., 1994a; Saikusa et al., 1994b). Consistent with these previous findings, the GABA content of Koshiguruma at 60 h germination and soaked in red onion solution was 153 mg/100 g (data not shown). In addition, germination in red onion solution resulted in up to a five times greater GABA content than that in water.
| Germination time | |||||
|---|---|---|---|---|---|
| 6 h after | 24 h after | 36 h after | 48 h after | ||
| Koshiguruma | Soaked in water | 5.2 | 18.0 | 30.8 | 49.4 |
| Soaked in red onion solution | 6.5(125%) | 30.4(169%) | 85.0(276%) | 123.9(251%) | |
| Koshihikari | Soaked in water | 3.6 | 7.7 | 12.3 | 15.9 |
| Soaked in red onion solution | 5.0(139%) | 13.9(181%) | 27.9(227%) | 76.9(484%) | |
| Hoshiyutaka | Soaked in water | 3.6 | 15.2 | 17.0 | 16.6 |
| Soaked in red onion solution | 3.5(97%) | 37.3(245%) | 61.4(361%) | 59.2(357%) | |
Units: mg/ 100g
A human study in which a fermented beverage containing 10–12 mg/100 mL of lactic acid bacteria-derived GABA was ingested daily for 12 weeks reported decreased systolic and diastolic blood pressure without alterations in heart rate (Inoue, 2003). Commercially-available germinated brown rice had germination times of 1–2 days, and the GABA content was generally 10–15 mg/100 g, similar to 8 mg/100 g in Koshihikari after 24 h germination in water in this study. On the other hand, the GABA content increased to 18 mg/100 g in Koshiguruma after 24 h germination in water and 30 mg/100 g in Koshiguruma after 24 h germination in red onion solution.
In addition, it was suggested that Koshiguruma contains more than twice the GABA content of Koshihikari and Hoshiyutaka at 24, 36, or 48 h germination; thus, it would be suitable as a functional rice material. It was reported that glucose metabolism improved when germinated brown rice was administered to high cholesterol-induced rabbits (Norhaizan et al., 2011). Moreover, isomalto-oligosaccharides used as prebiotics were found in germinated brown rice (Premsuda, 2008), and γ-tocopherol, γ-oryzanol, thiamine, niacin, and pyridoxine were also contained in germinated brown rice (Anuchita and Nattawat, 2010). Thus, it may be expected that germinated brown rice has activities besides lowering blood pressure.
Polyphenol content and DPPH radical scavenging capacity The polyphenol content and DPPH radical scavenging capacity of germinating samples are shown in Figure 3. The polyphenol content increased, but the DPPH radical scavenging capacity did not markedly change during germination. In addition, the polyphenol content and DPPH radical scavenging capacity were higher in samples germinated in red onion solution than in those germinated in water; however, this may be attributed to the presence of quercetin as a type of polyphenol.

Polyphenol content and DPPH radical scavenging capacity of germinated rice. shows germinated rice of Koshiguruma soaked in water, shows germinated rice of Koshiguruma soaked in red onion solution, shows germinated rice of Koshihikari soaked in water, shows germinated rice of Koshihikari soaked in red onion solution, shows germinated rice of Hoshiyutaka soaked in water, and shows germinated rice of Hoshiyutaka soaked in red onion solution. *: p < 0.05, *: p < 0.01: significantly different from each germinated rice at the same time.
Starch degrading enzymes To determine changes in endogenous enzymes during the germination of brown rice grains, we measured endogenous hydrolytic enzymes. During germination, α-amylase degrades starch to dextrin, leading to glucose production.
α-Amylase activity increased with germination time, while germination in red onion solution showed lower activity than germination in water at each period (Fig. 4). It has been reported that α-amylase activity decreased from day 3 of germination (Kiribuchi and Nakamura, 1974), and that germination rate and α-amylase activity are correlated (Phantipha et al., 2009, Premsuda et al., 2008). Although α-amylase activity increased during each period, its activity might decrease after day 3 of germination (Anuchita and Nattawat, 2010). There are reports that polyphenols inhibit saccharification enzymes such as amylase (Diganta et al., 2018; Oyedemi et al., 2017). Therefore, it is proposed that the quercetin contained in red onion was absorbed by the seeds during soaking and showed amylase inhibitory activity.

α-amylase activity during germination of rice cultivars. shows germinated rice of Koshiguruma soaked in water, shows germinated rice of Koshiguruma soaked in red onion solution, shows germinated rice of Koshihikari soaked in water, shows germinated rice of Koshihikari soaked in red onion solution, shows germinated rice of Hoshiyutaka soaked in water, and shows germinated rice of Hoshiyutaka soaked in red onion solution.
Cell wall degrading enzymes Cellulase activity decreased with germination, but was not affected by the red onion extract (Fig. 5). The activity of the cell-wall degrading enzyme cellulase did not increase during germination, and the effect of germination may vary depending on the kind of enzymes.

Cellulase activity during germination of rice cultivars. shows germinated rice of Koshiguruma soaked in water, shows germinated rice of Koshiguruma soaked in red onion solution, shows germinated rice of Koshihikari soaked in water, shows germinated rice of Koshihikari soaked in red onion solution, shows germinated rice of Hoshiyutaka soaked in water, and shows germinated rice of Hoshiyutaka soaked in red onion solution.
In this study, we evaluated metabolic changes associated with the germination of brown rice using the giant embryo rice cultivar Koshiguruma. As giant embryo rice is known to germinate slowly, we determined the most suitable pH and temperature conditions for Koshiguruma.
Germinated Koshiguruma brown rice showed a higher GABA content than either the germinated brown rice cultivar Koshihikari, a high-quality premium rice, or the high-amylose rice Hoshiyutaka. A red onion solution was employed for germination, and increases in the GABA content of samples were detected.
α-Amylase, a glycation-related enzyme, increased during germination, while no significant change was observed in the cell wall degrading enzyme cellulase. In addition, germination in the presence of red onion extract showed increased GABA and polyphenol contents as well as slightly increased antioxidant capacity.
Acknowledgements We would like to express our heartfelt thanks to the Niigata Crop Research Center and Kyushu University, who kindly provided the rice samples used in this study. The authors would like to thank Enago for the English language review.