マゲシカCervus nippon mageshimaeの遺伝的独自性についての再検討

論文ID: 2302
この記事には本公開記事があります。

詳細
論文ID: 2302
マゲシカ Cervus nippon mageshimae Kuroda and Okadaは、その形態的特徴によって記載されたニホンジカ C. nipponの亜種である。しかし、2014年の環境省のレッドデータブックの改訂に当たっては、亜種としての分布域が不明瞭であり、分類学的な位置づけが明確でないとされている(環境省 2014)。その一方で、近年実施された遺伝解析の結果からはマゲシカの遺伝的独自性が示されつつある。その遺伝的特徴から、マゲシカの分布域を明らかにし、近隣に生息するヤクシカ C. n. yakushimae(屋久島および口永良部島)やキュウシュウジカC. n. nippon(九州本土)との関係性を議論することが可能となってきた。そこで本研究では、マゲシカの遺伝的な特徴やその分布について、先行研究において報告されているミトコンドリアDNAのコントロール領域のデータを中心に再検討を行った。その結果、マゲシカの分布域とされる馬毛島および種子島の個体は、キュウシュウジカだけでなくヤクシカとも明確に異なる塩基配列を有していた。また核DNAやY染色体DNAに着目した先行研究には、マゲシカの生息地である種子島とヤクシカの生息地である屋久島間で対立遺伝子頻度に明確な違いがあることが示されていた。形態については、馬毛島および種子島のニホンジカと遺伝的にもっとも近いヤクシカとの間に明瞭な違いがあることが先行研究によって示されていた。一連の知見から、少なくとも馬毛島と種子島のニホンジカを九州に生息するニホンジカとは明確に異なる保全単位として認識することが適切であること、さらに2島間にもミトコンドリアハプロタイプの頻度をはじめとする集団遺伝学的な差異が存在する可能性が高いことが示唆された。
Cervus nippon mageshimae Kuroda and Okada (mage shika in Japanese) is a subspecies of C. nippon described based on its morphology. Although recent genetic studies indicate the genetic uniqueness of C. n. mageshimae, its description in the 2014 Red Data Book suggests that its distribution and taxonomic position as a subspecies need to be clarified. In this study, we re-evaluated the validity of C. n. mageshimae with genetic data collected in previous studies. Our phylogenetic analysis revealed distinct sequence differences in mitochondrial DNA between C. n. mageshimae and both C. n. nippon on Kyushu and C. n. yakushimae from Yaku-shima and Kuchinoerabu-jima Islands. Previous analyses of nuclear and Y-chromosome DNA have also identified differences in allele frequencies between the Tanegashima and Yaku-shima Island populations. In addition, previous morphological studies demonstrated morphological differences between these islands. Overall, these findings suggest the presence of genetically distinct populations on Mage-shima and Tanegashima Islands that can be recognised as conservation units.
ニホンジカ Cervus nippon は、偶蹄目シカ科に属する大型哺乳類であり、ロシア沿海州、中国、朝鮮半島、台湾、日本、ベトナムと東アジアを中心に広く分布している (大泰司 1986;Ohtaishi and Gao 1990)。日本列島においては北海道、本州、四国、九州、対馬列島、五島列島、大隅諸島、慶良間諸島などに分布している。これらの日本列島のニホンジカの亜種分類は、様々な学名・和名の変遷を経て (遠藤 1996)、大泰司 (1986) によって形態と分布情報から分類された6亜種:エゾシカ C. n. yesoensis、ホンシュウジカ C. n. centralis、キュウシュウジカ C. n. nippon、マゲシカ C. n. mageshimae、ヤクシカ C. n. yakushimae、ケラマジカ C. n. keramae が比較的広く受け入れられている。しかし1990年代以降のミトコンドリアDNA (mtDNA) や核DNAを用いた遺伝学的解析の結果 (例えばNagata et al. 1999;Goodman et al. 2001;Yamada et al. 2006;Nagata 2009) は、必ずしも従来の亜種分類とは一致しない。現在でも日本列島のニホンジカの種内分類群については議論があり(遠藤 1996)、保全や管理単位の設定における支障となっている。
マゲシカ Cervus nippon mageshimae Kuroda and Okadaは、馬毛島に生息するニホンジカの形態的特徴から、Kuroda and Okada (1950) によって記載されたニホンジカの亜種である。その後、馬毛島の個体群は、マゲシカ C. n. mageshimae (黒田 1953;今泉 1960) あるいはヤクシカ C. n. yakushimae (Imaizumi 1970; 今泉 1970; Corbet 1978; 日本哺乳類学会 1997) という和名および学名があてられてきた。大泰司(1986)によって馬毛島と種子島の個体群をマゲシカに含めること、ヤクシカとは別亜種にすることが提唱され、Tamate et al. (1998) や Goodman et al. (2001) は、種子島の個体群もCervus nippon mageshimae として扱っている。現在は両島以外に、鹿児島県の阿久根大島 (阿久根市誌編さん委員会 1974; Takatsuki 1980)、臥蛇島 (西之表市 1972;南日本新聞社 1981; 十島村誌編集委員会 1995)、喜界島 (永田ほか 2023) に人為的に移入された集団が生息する。しかし、2014年の環境省のレッドリストの改訂に当たっては、亜種としてのマゲシカの分布域が不明瞭であり分類学的な位置づけが明確でないという判断から、亜種マゲシカを認める立場をとらず、「馬毛島のニホンジカ」としてニホンジカC. nipponの地域個体群 (LP) としている (環境省 2014)。
近年実施された遺伝学的解析においてはマゲシカに一定の遺伝的独自性が存在し、共通の遺伝的特徴を有する集団が馬毛島および種子島に存在することが示されつつある。マゲシカの基準産地である馬毛島では、現在自衛隊基地の建設が計画されており、そのためのアセスメント事業において核マイクロサテライト (SSR) マーカーを用いた解析が実施されている (熊本防衛支局 2022)。その結果、種子島および馬毛島に生息する個体は、九州に生息するキュウシュウジカや屋久島に生息するヤクシカとは明瞭に異なっており、大泰司 (1986) や Goodman et al. (2001) で示されているマゲシカとするのが適切であるとしている (熊本防衛支局 2022)。また、マゲシカとしての言及ではないものの、種子島の個体を対象としたmtDNAや核SSRマーカー、Y染色体上のSSRマーカーを用いた解析において、キュウシュウジカおよびヤクシカとの遺伝的差異やそれらの生息地間の遺伝子流動の欠如を示すデータが示されている (例えば Terada et al. 2013;Yamashiro et al. 2015;Terada and Saitoh 2018;Takagi et al. 2022;永田ほか 2023)。
本研究では、マゲシカとされてきた馬毛島および種子島に生息するニホンジカの遺伝学的な独自性について再検討することを目的に、先行研究において報告されているmtDNAのコントロール領域のデータを集約し、再解析した。また、馬毛島および種子島間における集団遺伝学的な差異について検討するために、在来生息地である馬毛島および種子島間におけるmtDNAハプロタイプの組成と頻度についても比較した。そしてその解析結果および核DNAや形態に関する先行研究を参照しつつ、マゲシカの分布域とされてきた馬毛島および種子島に生息するニホンジカの遺伝的実態について考察した。
本研究における系統解析では、Yamada et al. (2007)、 Barančeková et al. (2012)、Terada et al. (2013)、 Yamashiro et al. (2015)、 永田ほか (2023) で公表されているニホンジカのmtDNAコントロール領域の全長塩基配列データを使用した (表1)。これらの先行研究で報告されている配列データのうち、九州地域に含まれる九州本土、対馬列島、五島列島、大隅諸島 (馬毛島、種子島、屋久島、口永良部島) のデータを解析に使用した。ただし、慶良間諸島のケラマジカは南九州に由来することが (Yamashiro et al. 2015)、喜界島のシカについては馬毛島に由来することが (永田ほか 2023)、共にmtDNAハプロタイプの解析から指摘されているため、これらの人為的な移入が明らかであるデータについては除外した。配列データはアメリカ国立医学図書館 (National Library of Medicine) のThe National Center for Biotechnology Information (https://www.ncbi.nlm.nih.gov/,2022年9月11日確認) より入手した。これらのデータについてMEGA11 (Tamura et al. 2021) に実装されているClustalW (Thompson et al. 1994) によりアライメントを行い、DnaSP v. 6. 12. 03 (Rozas et al. 2017) によりハプロタイプ判別を行った。
本研究で用いたすべてのハプロタイプ間の系統関係を検討するために分子系統樹を推定した。分子系統樹はMEGA11 (Tamura et al. 2021) により、近隣結合法 (Neighbor-Joining method,Saitou and Nei 1987) を用いて作成した。外群にはタイワンジカ C. n. taiouanus (アクセッションナンバー: AB279722) とアカシカ C. elaphus (NC_007704) を用いた。mtDNAコントロール領域中に含まれる繰り返し配列はNagata et al. (1999) を参考に識別し、除去した状態の851bpで分子系統樹を作製した。遺伝距離はKimura’s two-parameter method (Kimura 1980) を用い、ギャップについてはPairwise deletionオプションで算出した。統計学的有意性はブートストラップ法 (1,000回) で検証した。更に大隅諸島で確認されたハプロタイプ間の類縁関係を最節約ハプロタイプネットワーク図で表現した。ハプロタイプネットワークはTCS v1.2.1 (Clement et al. 2000) により推定した。大隅諸島のサンプルは繰り返し配列が全て4回であったため繰り返し配列は除去せず、mtDNAコントロール領域の全長配列をもとに解析を行った。ギャップは第5の塩基として扱い、連続するギャップについては注釈を加えたうえで1回の変異として作図した。
さらに馬毛島と種子島におけるmtDNAコントロール領域のハプロタイプの頻度について比較した。馬毛島のハプロタイプ頻度については、永田ほか (2023) に示されている5個体のデータを用いた。これらの5個体のデータは自然死した3個体のオス成獣と1個体の幼獣、1個体の胎仔から1996年5月に採取された筋肉サンプルを分析したもので、胎仔については死体で発見した雌成獣を解剖して取り出した個体である。種子島のハプロタイプ頻度は、2007年から2014年にかけて狩猟で捕獲された39個体の筋肉サンプルで決定されたハプロタイプから算出した。これらの内の10サンプルについてはTerada et al. (2013) に用いられている10サンプルと同一である。追加の29サンプルは、西之表市および中種子町において有害鳥獣捕獲された個体から得られたサンプルである。雌雄および推定年齢が記録されていた25個体については13個体がオス、12個体がメスであり、推定年齢は0歳が3個体、1歳が5個体、2歳以上が17個体であった。これらのサンプルのハプロタイプの決定もTerada et al. (2013) に準じて行った。これらの馬毛島と種子島のハプロタイプ多様度(h)と塩基多様度(π)、2島間の遺伝的分化を示すFST値はArlequin ver 3.5.2.2(Excoffier and Lischer 2010)を用いて算出した。
データベースから得られた995-1076塩基のmtDNAコントロール領域の全長配列に基づきハプロタイプを定義した結果、37のハプロタイプが確認された。ハプロタイプの命名は、大隅諸島のサンプルについては永田ほか (2023) に従い、それ以外のサンプルについてはそれぞれの先行研究 (Yamada et al. 2007;Barančeková et al. 2012;Terada et al. 2013;Yamashiro et al. 2015) で用いられているハプロタイプ名を踏襲した(表1)。また、mtDNAコントロール領域中に含まれる4から6回の繰り返し配列を除去した場合でも37ハプロタイプに分類されることに変わりなかった (表1)。近隣結合法を用いた分子系統樹では、マゲシカおよびヤクシカの分布域である大隅諸島から得られた8ハプロタイプ (Hap1-8) が、キュウシュウジカの分布域である九州本土や五島列島、対馬で確認された29ハプロタイプで構成されるクレードから区別され、95%のブートストラップ値で単系統群を構成した (表1、図1)。このうち、馬毛島、種子島で検出されたハプロタイプで構成されるマゲシカクレード (Hap1-5)と、屋久島、口永良部島で検出されたハプロタイプで構成されるヤクシカクレード (Hap6-8) が、それぞれ97%と80%のブートストラップ値で単系統群を構成した (図1)。
大隅諸島のサンプルを対象としたハプロタイプネットワークにおいても、マゲシカの分布域とされる馬毛島および種子島で検出されたハプロタイプと、ヤクシカの分布域である屋久島、口永良部島で検出されたハプロタイプ間に明確な塩基配列の違いがあることを明示した (図2a, b)。馬毛島および種子島で検出されたハプロタイプと屋久島および口永良部島で検出されたハプロタイプの間には、少なくとも7カ所の塩基置換 (Hap2とHap8) あるいは5カ所の塩基置換と7塩基の連続した挿入・欠失 (Hap2とHap6) が存在していた。馬毛島からはHap1および2が検出され、種子島からはHap2-5が検出された。Hap1およびHap3-5はいずれもHap2から派生していた (図2a)。
馬毛島と種子島間におけるハプロタイプの観察頻度には明瞭な違いが認められた (表2)。島間のFST値は有意にゼロより大きかった(FST = 0.479, p < 0.001 )。馬毛島における解析個体数は少ないものの、5個体中4個体から確認されたHap2は種子島においては39個体中の1個体のみの確認にとどまった。また馬毛島において確認されたHap1については、種子島からは確認されなかった (図2b)。一方で、種子島において比較的高頻度で確認されるHap3やHap5については、馬毛島からは確認されなかった。
マゲシカの遺伝的特異性およびレッドリストにおける評価について
今回の解析結果から、マゲシカの分布域とされる馬毛島および種子島の個体は、九州に生息するキュウシュウジカだけでなく、屋久島および口永良部島に生息するヤクシカとも明確に異なる塩基配列を有することが明らかになった。馬毛島と種子島から検出されたHap1-5は、屋久島をはじめとする他地域のハプロタイプとは明瞭に異なる塩基配列を有し、これらのシカが他の地域とは異なる独自の遺伝的特徴を持つことを示している(図1)。今回の系統解析およびハプロタイプネットワーク解析の結果は、馬毛島と種子島のシカが共通祖先を有することも示している。
本研究の結果は大泰司 (1986) によって提唱されている種子島と馬毛島に分布する亜種マゲシカの認識とも一致する。また、種子島と馬毛島においては、共通して検出されたHap2だけでなく、Hap1やHap3-5のような派生的なハプロタイプも検出された(図2)。キュウシュウジカやヤクシカが移入し種子島および馬毛島に小規模集団を維持していただけでは、系統間の変異の蓄積や系統内の派生的なハプロタイプの発生は生じない。したがって、種子島と馬毛島の系統は、変異の蓄積が可能な長期間にわたり孤立していた可能性、あるいは成立当初からある程度の変異を有する集団を起源とする可能性がある。
さらに一部の先行研究では、マゲシカ C. n. mageshimaeという亜種名は採用されていないものの、マゲシカに分類される種子島で採取された個体が解析に用いられている。これらの結果においても、以下に記述するようなマゲシカの遺伝的独自性やヤクシカに近縁という系統上の位置付けが示されている。そのうち、mtDNAの部分配列を用いた研究では、種子島と屋久島から採取された個体は九州本土や対馬、五島列島のニホンジカと比較して独自のクレードを形成することが明らかになっている (Yamada et al. 2007;Terada et al. 2013;Yamashiro et al. 2015)。さらに、Dong et al. (2022) によるミトコンドリアゲノムによる解析は、キュウシュウジカやホンシュウジカと、大隅諸島のシカが明確に区別されること、種子島の個体が屋久島の個体とはミトコンドリアゲノムのハプロタイプを共有せず、種子島と屋久島で異なるクレードを形成することを示している。本研究ならびに先行研究の一連の知見は、第3次レッドリストにおいてマゲシカについて不明とされた点が既に明らかになっていることを示している。不明瞭とされた分布域は基準産地である馬毛島と種子島であり、少なくとも遺伝的な特徴という点ではキュウシュウジカはもちろんヤクシカとも区別される。したがって、現在の「馬毛島のニホンジカLP」よりも「マゲシカの馬毛島個体群(LP)」とする方がより適切である。
日本列島に生息するニホンジカC. nipponの種内分類群についての見解は、大泰司 (1986) による6亜種:エゾシカ C. n. yesoensis、ホンシュウジカ C. n. centralis、キュウシュウジカ C. n. nippon、マゲシカ C. n. mageshimae、ヤクシカ C. n. yakushimae、ケラマジカ C. n. keramae が比較的広く受け入れられてきたものの、近年の遺伝解析はその問題点についても指摘してきた。ホンシュウジカとキュウシュウジカに相当する系統は認められないこと (Nagata et al. 1999;Goodman et al. 2001)、ケラマジカが人為的な移入に由来する可能性が高いこと (Tamate et al. 1998;Yamashiro et al. 2015) 等が示され、これらの種内分類群の見直しが必要であることは明らかである。また、大泰司 (1986) はホンシュウジカとしたものの、独立種C. pulchellus (Imaizumi 1970;環境庁 1991) あるいは亜種C. n. pulchellus (Corbet 1978) と扱われることもあった対馬に生息するツシマジカについても、大分県で捕獲されたキュウシュウジカと塩基配列が極めて近いハプロタイプが検出されていた(表1、Yamashiro et al. 2015)。そのためツシマジカについてはケラマジカ同様に人為的な移入を起源とする可能性を検討する必要がある。その一方で、ヤクシカとマゲシカについては、核SSR (Goodman et al. 2001) やミトコンドリアゲノム (Dong et al. 2022) を用いた先行研究においても、キュウシュウジカとは異なるクレードを形成し、屋久島と種子島間でハプロタイプを共有しない等、遺伝的な独自性が確認されており、他の地域に生息するニホンジカとは異なる進化的背景を有していることが明らかになった。
馬毛島および種子島間のハプロタイプ頻度の違いと逸出個体の起源推定への利用可能性
馬毛島および種子島間のハプロタイプ頻度の違いは、2島間の個体の移動が制限されていることを示唆する。今回の結果は、馬毛島における解析個体が5個体と少ないものの、馬毛島および種子島間にハプロタイプの組成および頻度に明瞭な違いがあることを示している (表2)。馬毛島および種子島の集団は、近年においても大きな個体群サイズの変動を経験しており、集団内のハプロタイプ頻度が遺伝的浮動によって変化した可能性はある。馬毛島、種子島ともに戦前、戦後の一時期に絶滅寸前、あるいは数十頭程度まで個体数を減らしたという記述があり (西之表市編纂委員会 1971;下野1982)、その後、馬毛島では700〜1000頭程度まで、種子島では6000頭程度まで個体数が回復している (鹿児島県 2022;熊本防衛支局 2022)。しかしながら、ハプロタイプの頻度だけでなく、その種類も異なっていたことから、馬毛島および種子島間の遺伝子流動が長期間制限されていたと考えられる。また、両島で共通して確認されたHap2が、ハプロタイプネットワークの基部に位置するのに対し、馬毛島でのみ確認されたHap1、種子島のみで確認されたHap3、Hap4、およびHap5は、それぞれHap2から派生したハプロタイプとなっている(図2)。したがって、これらの島固有のハプロタイプはそれぞれの島で発生した可能性が高い。
このような馬毛島および種子島におけるハプロタイプ頻度の違いは、人為的に導入されたマゲシカの由来の解明にも重要な情報を提供する。マゲシカは、鹿児島県の阿久根大島 (阿久根市誌編さん委員会 1974;Takatsuki 1980)、臥蛇島 (西之表市 1972;南日本新聞社 1981;十島村誌編集委員会 1995)、喜界島 (永田ほか 2023) に導入され、現在でも生息が確認されている。さらに、福岡県能古島、鹿児島県長島などにも馬毛島のシカが導入されていた記録がある (安部 1926;早川 1942;西之表市 1983)。喜界島では馬毛島のみで確認されているHap1だけが検出されており、喜界島に導入された個体の起源が馬毛島であることが示唆されている (永田ほか 2023)。また、馬毛島のマゲシカとして飼育されている鹿児島県鹿児島市の平川動物園の個体からもmtDNAコントロール領域の部分配列(全長998bp中694bp)がHap1と一致することが確認されており (Terada and Saitoh 2018, AB771718)、この飼育個体が馬毛島由来であることを裏付けている。Hap1は、永田ほか (2023) で分析された馬毛島の5個体中の1個体からしか確認されていない。しかし、導入された喜界島や平川動物園の個体においてもHap1が確認されていることから、馬毛島においてある程度の頻度で存在していた可能性が高い。Hap2については、神奈川県の丹沢の野生個体からも確認されており (Hata et al. 2018;Hata et al. 2019)、馬毛島あるいは種子島から人為的に導入され逸出した個体を起源とすると考えられる。
人為的導入と逸出が遺伝解析により判明した例は神奈川県の丹沢の事例だけではない。人為導入されたとみられるヤクシカC. n. yakushimaeが野生化し、その遺伝子が在来のホンシュウジカC. n. centralisと集団内に拡散している事例が存在することも明らかとなっている (Eva and Yamazaki 2018;Yamazaki 2018)。mtDNAの分析では、富山県内の個体のおよそ40%がヤクシカ由来と推定されるハプロタイプを有しており(Yamazaki 2018)、人為的な導入と逸出が遺伝構造を大きく攪乱する可能性を示唆している。このほかにも北海道(Terada et al. 2013)や南関東(Yuasa et al. 2007)において人為的な導入と逸出に由来すると考えられるハプロタイプが野生個体から検出されている。遺伝構造の攪乱の現状を把握するためにも、その起源を推定するためにも、ニホンジカの地域個体群における遺伝構造やその系統的特徴の把握は重要である。
大隅諸島間および九州間での遺伝子流動の有無
ニホンジカのmtDNAによる系統解析については比較的多くのデータが公開されている一方で、核DNAを用いた集団遺伝学的研究、特にマゲシカの分布域である馬毛島と種子島と、ヤクシカの分布域である屋久島と口永良部島に着目した研究は限られている (Goodman et al. 2001;Terada and Saitoh 2018)。しかしながら、核SSRマーカーやY染色体上のSSRマーカーによる先行研究は、九州と大隅諸島間はもちろん、種子島と屋久島間での個体の移動が明確に制限されていることを示している。核SSRマーカーによって解析しているGoodman et al. (2001) およびTerada and Saitoh (2018)、Y染色体上のYSSRマーカーによって解析しているTakagi et al. (2022) は、九州と大隅諸島(種子島・屋久島)間で明確に対立遺伝子頻度に違いがあることを示しており、両地域間の個体の移動が極めて限定的であることを示唆している。さらにTerada and Saitoh (2018) は核SSRマーカーの遺伝子型データから種子島と屋久島間の遺伝的分化の指標であるFST値を0.32と報告しており、両島間での遺伝子流動が限定的であることが示されている。また、雄の移動の指標となると考えられるYSSRマーカーによる解析では、種子島と屋久島間での共通のハプロタイプは検出されていない (Takagi et al. 2022)。外部形態の比較においても種子島、屋久島間での違いが報告されている (Terada et al. 2012;寺田・齊藤 2014;Terada and Saitoh 2018)。これらの結果から、種子島と屋久島間における個体の移動と移入後の繁殖成功、すなわち遺伝子流動が制限されていたことは明らかである。
マゲシカの人為移入起源説の否定と進化史考察における重要性
マゲシカはKuroda and Okada (1950) によって、キュウシュウジカとヤクシカの中間的な特徴を有すると記載されている。おそらくそのような形態的な特徴から、過去には、馬毛島あるいは種子島の個体が移入に由来するという考察もされてきた (今泉 1970;今泉・小原 1976;松元ほか 1984)。東ヨーロッパのアカシカC. elaphusを対象とした遺伝解析の結果は、中世以降のアカシカの人為的な移動と再導入の両方が、その遺伝的特徴に強い影響を及ぼしていることを示し (Niedziałkowska a et al. 2011)、古代DNA (Ancient DNA) の解析は新石器時代に人間がアカシカをスコットランドに持ち込んだ可能性を指摘している (Stanton et al. 2016)。ニホンジカにおいても、奈良県の春日山周辺の個体群が1000年以上にわたって人為的に保全され残存したものであることが示唆されている (Takagi et al. 2023)。これらの事例や先述したケラマジカやツシマジカの事例を踏まえれば、ニホンジカの種内系統や分布の形成過程を考察する際には、人為的な影響を十分に考慮する必要がある。しかしながら、本研究で整理した遺伝解析の結果からは、人為的な移入であれば確認されるはずの同一ハプロタイプあるいは遺伝的に近縁なハプロタイプが他地域で確認されず、馬毛島および種子島に生息するマゲシカが独自の系統であることが示された(図1、図2)。したがって、馬毛島および種子島に生息するマゲシカは人為的な移入に由来するという説は明確に否定される。
マゲシカと他の亜種との関係においては、分子系統解析の見解と形態学的な見解は必ずしも一致しない。分子系統解析において、マゲシカはキュウシュウジカよりもヤクシカに近いことが明確に示されてきた (図1、Goodman et al. 2001;Yamada et al. 2007;Terada and Saitoh 2018;Takagi et al. 2022;永田ほか2023)。その一方で、形態学的な見解においては、マゲシカとヤクシカとの違いが明確にされており、むしろキュウシュウジカとの類似性が示されている (松元ほか 1984;城間・太田 1996;Terada et al. 2012;Terada and Saitoh 2018)。共通祖先を有するにも関わらずマゲシカとヤクシカが異なる形態を有するのは、島嶼における孤立とそれぞれの生息環境へ適応による可能性が高い (Terada and Saitoh 2018)。したがって、両亜種を対象とした今後のさらなる遺伝的、形態的比較研究は、大型哺乳類の島嶼適応を理解するうえで重要な知見を提供すると期待できる。現在の地形と海峡間の個体の移動にだけ着目するのではなく、海水面の変動に伴う島嶼の連結や分断を考慮に入れつつ、それぞれの島嶼の集団の形成史について、今後検討していく必要がある。
マゲシカの保全管理上の提言
本研究では、馬毛島と種子島に独自の遺伝的特徴を有する集団が成立していることが示された。この2つの島においてどのように系統が維持されてきたのか、環境や地理的要因を明らかにするとともに、両生息地において適切に保全管理を実施し、系統のみならず進化プロセスの背景ごと保全することが重要である。一方で、現在馬毛島では、自衛隊基地の建設が計画されており (熊本防衛支局 2022)、島面積の61%を改変する大規模な開発が予定されている。2つの島に限られるマゲシカの生息地のひとつにおいて生息状況に大規模な変化が生じる事態であり、科学的な見地から馬毛島個体群を保全するあらゆる取り組みを講ずるべきである。
本研究ではさらに、馬毛島と種子島間におけるmtDNAハプロタイプの組成の明瞭な違いが示された。今後、核DNAを対象とした解析により、両島の集団の遺伝的な違いがより明確にされると期待される。また、種子島のシカをCervus sp. とし、馬毛島のマゲシカと区別して形態を分析している松元ほか (1984) は、大きさを表すとする第一主成分において種子島のCervus sp.は雌雄双方でマゲシカよりも大きいことを、頭蓋骨形態の主成分分析によって示している。したがって、マゲシカとヤクシカ、キュウシュウジカの地域個体群を網羅した形態に関する比較研究やその特徴の整理は、この地域のシカにおける島嶼適応を知るうえでも今後の重要な課題である。一方、自衛隊基地建設に際してのアセスメント事業においては、核SSRマーカーを用いた解析の結果としてマゲシカの遺伝的特異性を認める一方で、“馬毛島の個体群に固有の遺伝子は確認されませんでした”としている (熊本防衛支局 2022)。しかしながら、上述のmtDNAについて検討していないほか、対立遺伝子の固有性に関する議論についても疑問が残る。対立遺伝子の比較対象には馬毛島の個体が人為的に導入された阿久根大島が含まれている。分析された核SSR遺伝子座の遺伝子型データが公開されていないために断定はできないものの、馬毛島に固有の対立遺伝子が存在し、それが阿久根大島にも受け継がれているために、馬毛島個体群として固有の対立遺伝子が無いようにみえている可能性も残されている。つまり、在来生息地としては馬毛島にしか存在しない対立遺伝子も存在しうる。残念ながら、2001年以降、馬毛島における調査やサンプリングが認められない状況にある (立澤 2021)。その制約が解消され、今後馬毛島の個体を含めた網羅的な研究を進めることができれば、マゲシカの実態をより正確に把握することができるであろう。このことはさらに、大隅諸島のニホンジカの由来、ひいては日本列島のニホンジカの集団構造の全容解明に貢献し、適切な保全や管理単位の設定が可能となるだろう。
謝 辞
Richard P. Shefferson氏には英文の校閲をしていただいた。本研究は、日本学術振興会科学研究費・基盤研究 (A)(20H00652) による成果である。
本研究で使用したニホンジカのミトコンドリアDNAコントロール領域ハプロタイプ
| ハプロタイプ* | アクセッション ナンバー ** | 原著論文中の ハプロタイプ名 | サンプル採取地域 | 亜種名† | 引用文献 |
|---|---|---|---|---|---|
| Hap1 | LC712850 | Hap 1 | 鹿児島県(馬毛島) | マゲシカ | 永田ほか 2023 |
| Hap2 | AB757723 | Tng3(4) | 鹿児島県(種子島) | マゲシカ | Terada et al. 2013 |
| Hap2 | LC712848, LC712849, LC712851, LC712852 | Hap 2 | 鹿児島県(馬毛島) | マゲシカ | 永田ほか 2023 |
| Hap3 | AB279718 | 4Tng1 | 鹿児島県(種子島) | マゲシカ | Yamada et al. 2007 |
| Hap4 | AB757724 | Tng4(4) | 鹿児島県(種子島) | マゲシカ | Terada et al. 2013 |
| Hap5 | AB279719 | 4Tna2 | 鹿児島県(種子島) | マゲシカ | Yamada et al. 2007 |
| Hap6 | AB279721 | 4Yks2 | 鹿児島県(屋久島) | ヤクシカ | Yamada et al. 2007 |
| Hap7 | AB279720 | 4Yks1 | 鹿児島県(屋久島) | ヤクシカ | Yamada et al. 2007 |
| Hap8 | LC712853–LC712857 | Hap 8 | 鹿児島県(口永良部島) | ヤクシカ | 永田ほか 2023 |
| 4Fko1/KY4E | AB279714 | 4Fko1 | 福岡県 | キュウシュウジカ | Yamada et al. 2007 |
| 4Fko1/KY4E | No Number | KY4E | 大分県 | キュウシュウジカ | Yamashiro et al. 2015 |
| 4Gto1/KY4L/J824 | AB279717 | 4Gto1 | 長崎県(五島列島) | キュウシュウジカ | Yamada et al. 2007 |
| 4Gto1/KY4L/J824 | JF893526 | J824 | 長崎県 | キュウシュウジカ | Barančeková et al. 2012 |
| 4Gto1/KY4L/J824 | No Number | KY4L | 長崎県 | キュウシュウジカ | Yamashiro et al. 2015 |
| 4Stm1 | AB279715 | 4Stm1 | 鹿児島県 | キュウシュウジカ | Yamada et al. 2007 |
| 4Tsm1 | AB279716 | 4Tsm1 | 長崎県(対馬) | キュウシュウジカ | Yamada et al. 2007 |
| 5Myz1/KY5C | AB186348 | 5Myz1 | 宮崎県 | キュウシュウジカ | Yamada et al. 2007 |
| 5Myz1/KY5C | AB871980 | KY5C | 宮崎県,鹿児島県 | キュウシュウジカ | Yamashiro et al. 2015 |
| 5Myz2 | AB279723 | 5Myz2 | 宮崎県 | キュウシュウジカ | Yamada et al. 2007 |
| 5Myz3 | AB279724 | 5Myz3 | 宮崎県 | キュウシュウジカ | Yamada et al. 2007 |
| 5Myz4 | AB279725 | 5Myz4 | 宮崎県 | キュウシュウジカ | Yamada et al. 2007 |
| KY4A | AB871968 | KY4A | 宮崎県,鹿児島県 | キュウシュウジカ | Yamashiro et al. 2015 |
| KY4B | AB871969 | KY4B | 熊本県 | キュウシュウジカ | Yamashiro et al. 2015 |
| KY4C | AB871970 | KY4C | 大分県 | キュウシュウジカ | Yamashiro et al. 2015 |
| KY4D | AB871971 | KY4D | 大分県 | キュウシュウジカ | Yamashiro et al. 2015 |
| KY4F | AB871972 | KY4F | 大分県 | キュウシュウジカ | Yamashiro et al. 2015 |
| KY4G | AB871973 | KY4G | 大分県 | キュウシュウジカ | Yamashiro et al. 2015 |
| KY4H | AB871974 | KY4H | 大分県 | キュウシュウジカ | Yamashiro et al. 2015 |
| KY4I | AB871975 | KY4I | 大分県 | キュウシュウジカ | Yamashiro et al. 2015 |
| KY4J | AB871976 | KY4J | 大分県 | キュウシュウジカ | Yamashiro et al. 2015 |
| KY4K | AB871977 | KY4K | 大分県 | キュウシュウジカ | Yamashiro et al. 2015 |
| KY5A | AB871978 | KY5A | 鹿児島県 | キュウシュウジカ | Yamashiro et al. 2015 |
| KY5B | AB871979 | KY5B | 鹿児島県 | キュウシュウジカ | Yamashiro et al. 2015 |
| KY5D | AB871981 | KY5D | 熊本県 | キュウシュウジカ | Yamashiro et al. 2015 |
| KY5E | AB871982 | KY5E | 大分県 | キュウシュウジカ | Yamashiro et al. 2015 |
| KY5F | AB871983 | KY5F | 大分県 | キュウシュウジカ | Yamashiro et al. 2015 |
| KY5G | AB871984 | KY5G | 大分県 | キュウシュウジカ | Yamashiro et al. 2015 |
| KY5H | AB871985 | KY5H | 大分県 | キュウシュウジカ | Yamashiro et al. 2015 |
| KY5I | AB871986 | KY5I | 大分県 | キュウシュウジカ | Yamashiro et al. 2015 |
| KY5J | AB871987 | KY5J | 大分県 | キュウシュウジカ | Yamashiro et al. 2015 |
| KY6A | AB871988 | KY6A | 鹿児島県 | キュウシュウジカ | Yamashiro et al. 2015 |
| KY6B | AB871989 | KY6B | 熊本県 | キュウシュウジカ | Yamashiro et al. 2015 |
* 反復配列を除去したハプロタイプであり、図1に対応する。
**KY4Eと KY4Lはそれぞれ,4Fko1 (AB279714) と4Gto1 (AB279717)と同一であることがYamashiro et al.(2015)に記載されているものの、データベースに配列登録が行われていないためNo Numberとしている。
†亜種分類は大泰司 (1986)に従った。
馬毛島および種子島におけるミトコンドリアDNAコントロール領域のハプロタイプ多様度、塩基多様度およびハプロタイプ頻度
|
n |
ハプロタイプ多様度h | 塩基多様度π | ハプロタイプ* | |||||
|---|---|---|---|---|---|---|---|---|
| Hap1 | Hap2 | Hap3 | Hap4 | Hap5 | ||||
| 馬毛島 | 5 | 0.400 | 0.0008 | 1 | 4 | 0 | 0 | 0 |
| 種子島 | 39 | 0.549 | 0.0007 | 0 | 1 | 24 | 3 | 11 |
*本研究において定義したハプロタイプ名。先行研究におけるハプロタイプ名との対応は表1を参照。
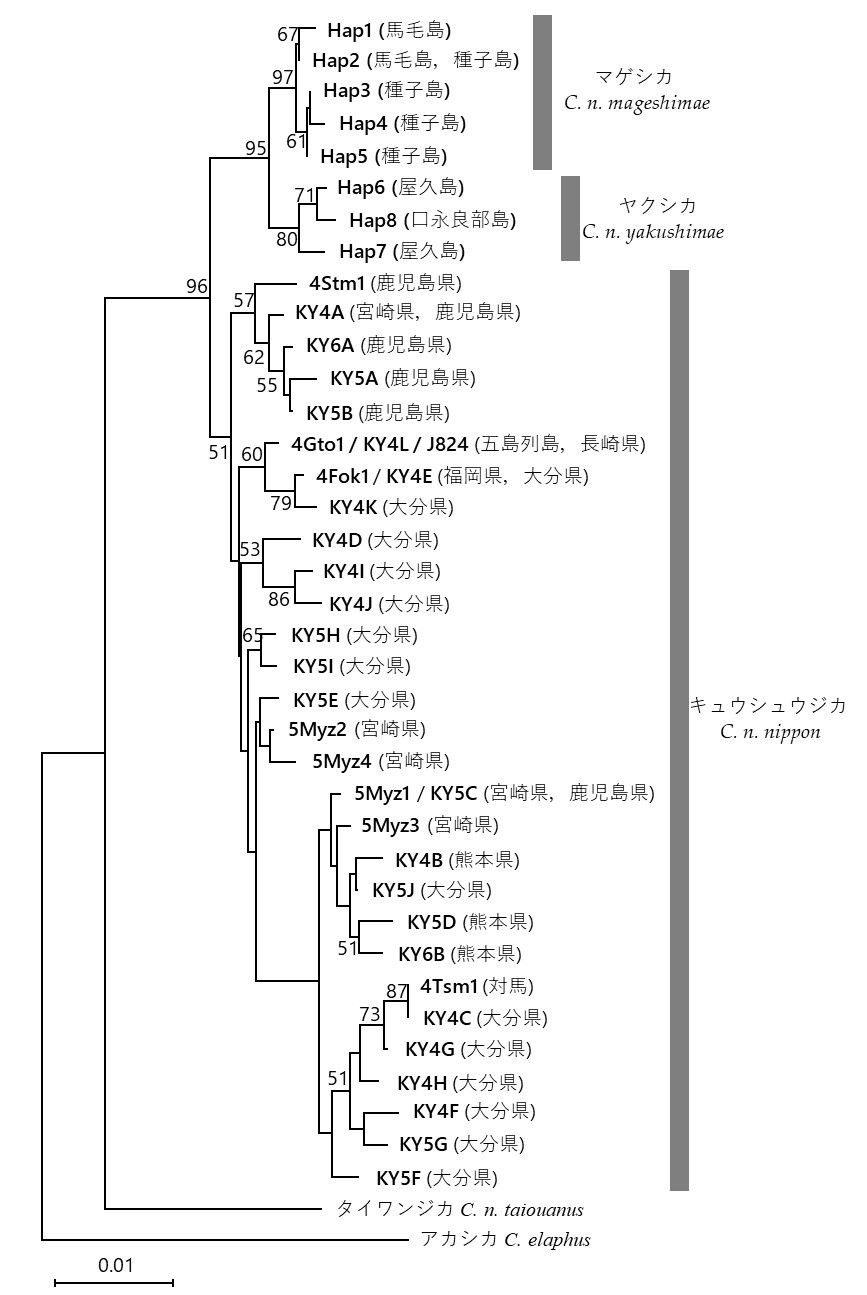
ニホンジカのミトコンドリアDNAコントロール領域の近隣結合法(Neighbor-Joining method)による分子系統樹。スケールバーは遺伝距離を表し、ブートストラップ値が50%以上の分岐点にはその数値を示した。ハプロタイプの詳細は表1を参照のこと。

(a)大隅諸島(馬毛島、種子島、屋久島、口永良部島)のニホンジカのミトコンドリアDNAコントロール領域の最節約ハプロタイプネットワーク。黒い太線はハプロタイプ間の一塩基置換を、白い太線は挿入・欠失を、白い丸は未検出ハプロタイプを表している。なおアスタリスク(*)の付いている白い太線は連続する7つの挿入・欠失を表す。(b)大隅諸島の地図と各島嶼におけるハプロタイプの分布。ハプロタイプの詳細は表1を参照のこと。