イトウは国際自然保護連合(IUCN)のレッドリストにCRとして掲載されている国内最大級の淡水魚であり、土地開発があまり進んでいない湿原や湿地帯をその流域に含む河川に生息する。そのため、湿地帯に形成される河川の氾濫原はイトウの生息環境に重要であると考えられてきた。しかし、本種に関するこれまでの知見は成魚に関するものが多く、幼魚に関する科学的知見は乏しい。本研究では、イトウの幼魚から成魚までの生息環境特性を明らかにするとともに、同所的に生息する同科魚類との比較を行い、本種の保全対策に寄与することを目的とした。調査は北海道大学雨龍研究林を流れるブトカマベツ川で行った。本河川には氾濫原が存在し、川筋が幾本にも分かれる網状流路が発達している。調査は網状流路が形成する分流域と本流域の2つに分けて実施し、河川規模の小さい分流域ではエレクトロフィッシャーを用いた捕獲を行い、河川規模の大きい本流域ではシュノーケリングを用いた潜水目視を行った。さらに、調査地点の物理環境と捕獲された個体の胃内容物を調べた。30地点で実施した分流域調査の結果、捕獲されたイトウは尾叉長69-137mmの幼魚であった。分流域の物理環境について主成分分析を行った結果、流速が遅く濁度が高いという止水的環境においてイトウ幼魚の生息密度が高くなる傾向が認められた。イトウ幼魚の胃内容物からは、魚類や両生類といった大型動物や動物プランクトンのミジンコ目が確認され、イワナおよびヤマメと比べて陸生落下動物の割合が少なかった。21地点で行った本流域調査の結果、目視されたイトウはいずれも体長300-800 mmの若魚・成魚であった。本流域の物理環境の主成分分析の結果、倒木などのカバー割合が高く深い淵においてイトウ若魚・成魚の生息密度が高くなる傾向が認められた。以上の結果から、イトウ幼魚は氾濫原に形成される流速が極めて遅い場所を選択的に利用するのに対し、イトウ若魚・成魚は流れのある本流で深くカバーのある環境を選択的に利用し生息していることが明らかになった。また、イトウ幼魚は成魚と同様に魚食性を示すことに加え、他のサケ科魚類が選好する陸生落下昆虫以外の餌資源を多く利用することが分かり、イトウは幼魚のときから他のサケ科魚類とは異なる摂餌行動をもつと考えられた。今後、イトウの野生個体群を保全していくためには、氾濫原環境の保全が極めて重要であると考えられた。
The Sakhalin taimen Parahucho perryi, one of the largest freshwater fish in Japan, is listed as Critically Endangered (CR) on the IUCN Red List. It occurs in rivers that contain marshes and wetlands with little land development. Therefore, floodplains are considered essential habitat for this species. However, most of our knowledge of Sakhalin taimen ecology concerns adults, and there is limited scientific knowledge of juveniles. In this study, we sought to identify the habitat characteristics of juvenile and adult Sakhalin taimen and compare them with those of sympatric fish species. The study took place in the Butokamabetsu River, which flows through the Uryu Experimental Forest of Hokkaido University and has a floodplain and well-developed anastomosing channels. Two areas were surveyed: the secondary side-channels, via electrofishing, and the main channel, via snorkelling. We also examined the physical environment of the study sites and the stomach contents of captured fishes. Thirty side-channel sites were surveyed, and all captured Sakhalin taimen were juveniles with fork lengths of 69–137 mm. Principal component analysis of the physical environment showed that juvenile Sakhalin taimen tended to be more abundant in lentic environments with low flow velocities and high turbidity. The stomachs of juvenile Sakhalin taimen contained fish, amphibians, and zooplankton (Cladocera), and a smaller proportion of terrestrial insects compared to the sympatric white-spotted charr Salvelinus leucomaenis and masu salmon Oncorhynchus masou. Twenty-one main-channel sites were surveyed, and all observed Sakhalin taimen were subadults or adults with fork lengths of 300–800 mm. Principal component analysis of the physical environment showed that subadult/adult Sakhalin taimen tended to be more abundant in deeper pools with more cover structure, such as woody debris. These results indicate that juvenile Sakhalin taimen select side channels with very slow currents, whereas subadult/adult Sakhalin taimen use deep pools in the main channel with sufficient cover. Conservation of floodplains and anastomosing channels is vital for the persistence of Sakhalin taimen in the wild.
イトウParahucho perryiはサケ科に分類される国内最大級の淡水魚で、成魚の体長は1 m以上、体重は25-45 kgに達する(福島ほか 2008)。本種は国際自然保護連合のレッドリストでは深刻な危機(CR)(IUCN 2022)、環境省のレッドデータブックに絶滅危惧IB類(EN)(環境省 2020)に指定されており個体群の保全が求められている。現在日本国内の北海道本島に7つの安定個体群の存在が知られているほか、択捉島、サハリン島およびロシア極東地域の日本海沿岸にも分布する(江戸 2007; 福島 2015; 大泰司2017)。昭和初期までは青森県小川原湖にも生息したが、乱獲により絶滅に追いやられたといわれる(日下部1988)。イトウは多数回繁殖を行い、春季に河川上流部の淵尻に産卵することが知られており、在来サケ科魚類で春季に産卵を行うのはイトウのみである(福島ほか 2008)。7月から8月にかけ体長3 cm程の稚魚が産卵床から浮上し、翌年には7 cm程度になり、20 cm以上に成長するのに3年かかるとされている(山代 1965, 1983; 久保 1990)。また、イトウは高い母川回帰性をもつと考えられており(江戸・東2002; Fukushima and Rand 2021)、朱鞠内湖の流入河川では数km離れた河川間においても遺伝的分化が認められている(畑山ほか 2017)。イトウは長年にわたりHucho属の1種とされてきたが、本種は海水適応能力をもち(久保 1989)、降海型が広範囲に存在する(Arai et al. 2004; Honda et al. 2010; Suzuki et al. 2011)といった他のHucho属とは異なる生態的特性をもち、さらに遺伝的な差異も大きいことから、現在はサケ科に属する1属1種と考えられている(福島 2015)。
イトウが分布する流域は、湿地や海跡湖の分布と対応する傾向にあることが知られている(福島ほか 2008)。そのため、イトウの生息には湿地や潟湖の存在が必要であり、1960年代後半から1970年代にかけての土地改良事業や農地面積の増大にともないイトウの絶滅が進行したと指摘されている(鷹見・川村 2008; Fukushima et al. 2011)。とくに氾濫原は稚魚期のイトウの生息環境として必要であると指摘されているが(江戸・東 2002; 江戸 2007)、農地化や土地開発、護岸工事など河川改修による河川の直線化によって氾濫頻度が減り、氾濫原環境が減少してきた。氾濫原に依存した生活史をもつイタセンパラAcheilognathus longipinnisやアユモドキParabotia curtusなど本州に生息する希少淡水魚も個体群の絶滅が進行し、天然記念物や絶滅危惧種に指定されるに至っている(小川 2008; 永山 2013; 渡辺 2016)。イトウ幼魚が選択的に利用しているであろう氾濫原特異的な生息環境を明らかにすることは、イトウの個体群存続性を左右する一つの要因の特定化に繋がると期待される。しかし、イトウ幼魚の生息環境や食性に関する知見は限られており(江戸 2007; 江戸・東 2002;佐川ほか 2003; 鈴木 2016)、とくに生息密度と物理環境の関係や生息場所と非生息場所の物理環境の相違についてはこれまで定量的なデータは示されていない。
これまでのイトウの生息場所に関する知見は、本種の特色が大型であるということや、繁殖場所の保全の重要性などから、成魚に関するものが多い(森ほか 1997; Fukushima 2001; 佐川ほか 2002; 本多 2020)。イトウの生活史で最も死亡率の高い稚魚、幼魚期の生態、行動は情報が少なく、今後、保護対策を検討するためには、幼魚に関する定量的な科学的知見の蓄積が必要である(福島 2015)。本研究では、イトウの幼魚に関する生態学的知見を得ることに焦点をあて、夏季の生息環境の特性を調べて、イトウの成魚と幼魚の生息環境の特性比較、イトウ幼魚の餌生物の把握と同所的に生息する同科魚類のヤマメOncorhynchus masouおよびイワナSalvelinus leucomaenisと比較し、本種の保全対策検討に寄与することを目的とした。
調査地
本研究は北海道北部に位置する朱鞠内湖の流入河川であるブトカマベツ川で行った(図1)。朱鞠内湖は1943年の雨竜ダム建設に伴い完成した人造湖であり、日本最大の23.73 km2の湛水面積をもつ。ブトカマベツ川は資源保護のために全魚種禁漁の措置が講じられているほか、流域の大部分が北海道大学雨龍研究林内を流れるため一般のアクセスは制限されている。比較的高い自然度を保つブトカマベツ川は、平水時の河道から氾濫して冠水する場所である氾濫原が存在し、川筋が幾本にも分かれる網状流路の形態が発達している。氾濫原のなかで代表的なものに旧河道があり、本流から分岐しているもの、本流とは通常時は接続せず増水時のみ接続するもの、本流と一切接続しないものがあり、プランクトンや魚類、両生類などの生息地を提供している(Uno et al. 2022)。本研究では、網状流路を構成する分流域と本流域において、異なる手法を用いてフィールド調査した。
分流域の調査
2021年7月から9月にブトカマベツ川分流域で、任意に選んだ30か所の淵を調査地点と設定した(平均値 ± SD, 長さ10.0 ± 4.8 m)。地点ごとにエレクトロフィッシャー(300-400 V DC, model 12B, Smith-Root, Inc., Vancouver, WA, USA)とたも網を用いて3回の魚類捕獲を行った。捕獲されたイトウ、イワナ、ヤマメの3種のサケ科魚類について、フェノキシエタノール(約0.5 ml/L)で麻酔を施した後、個体ごとに尾叉長と体重を記録した。3回の採捕数のデータからプログラムCAPTUREのModel 2(mbh-Pollock法)を用いた除去法により地点内の魚種別個体数を推定し(Katano 2010)、水面面積あたりの生息密度を求めた。
各調査地点の物理環境として、水温、濁度、川幅、水深、流速、底質粒径、河岸のえぐれや倒木により形成されたカバー割合を計測し記録した(付録1 図1)。水温は調査地点ごとに流程を4等分する縦断測線の中央3点で計測し、平均値を算出するとともに、計測時刻を記録した。また、ブトカマベツ川本流の泥川横断林道の橋付近の3地点で自動記録水温計(StowAway TidbiT, Onset社)を用いて15分おきに水温を計測し、時刻毎に3地点の平均値を算出し基準水温として扱い、調査地点の調査時刻の水温と基準水温の偏差を調査地点の水温偏差として算出し、時間的変化の大きい水温環境を評価した。濁度は濁度計(YSI ProDSS, ワイエスアイ・ナノテック社)を用いて流程を4等分する縦断測線の中央点3点で計測し平均値を算出した。川幅は地点内に設定した1 m間隔の横断測線をレーザー距離計(DISTO D3, Leica社)により0.1 m単位で計測した、地点内の平均値を算出した。水深は1 m間隔の横断測線をさらに4等分する3点で、直尺を用いて最小単位1 cmで計測し、地点内の平均値を算出した。流速は流速計(VR-301, ケネック社)を用い、水深と同じ計測点で、水深の6割深度の10秒平均流速を最小単位0.1 cm/sで計測し、地点内の平均値を算出した。底質は水深と同じ計測点で採取し、泥、砂、礫を目視判別し、泥は一律0.00395 cm、砂は一律0.1 cm、礫は主たる礫の短軸を最小単位1 cmで実測し、地点内の底質粒径の平均値を算出した。カバー割合は、河岸のえぐれおよび倒木を最小単位0.1 mで縦横を計測し、長方形または台形で近似し、地点ごとに水面面積に対するカバー面積の割合として算出した。
本流域の調査
2022年8月にブトカマベツ川本流域で、任意に選んだ21か所の淵を調査地点と設定した(平均値 ± SD, 長さ16.4 ± 7.9 m)。地点ごとにシュノーケリングによる潜水調査を行い、地点内で目視されたイトウの個体数と体長(100 mm単位)を記録した。他のサケ科魚類については概数を記録した。目視されたイトウの個体数と水面面積を用い生息密度を推定した。なお、本流域にはエレクトロフィッシャーを用いた魚類の捕獲が困難な深い淵が多く存在するため、潜水目視調査法を用いた。また、各調査地点の物理環境として、川幅、水深、流速、底質粒径、カバー割合を計測した。川幅は各地点内に5本以上の横断測線を等間隔に設定しレーザー距離計(GLM 50-23G, Bosch社)を用いて0.1 m単位で計測し平均値を算出した。水深、流速、底質粒径は、川幅を計測した横断測線を4等分する3点で、分流調査時と同様の方法で計測・算出した。カバー割合についても分流調査時と同様の方法で計測・算出した。流れのある本流の水温および濁度は地点間の差が小さいと考えられたため計測しなかった。
成長段階の区分
佐川(2006)は、全長>400mmは成魚、200-400mmは若魚(>2+歳)、90-200mmは幼魚(1+歳)、<90mmは稚魚(0+歳)とイトウの成長段階を区分している。本研究の分流域調査で捕獲されたイトウは尾叉長が69-137 mmであり(後述)、山代(1965)および久保(1990)を参考に年齢を推定すると1+歳魚もしくは2+歳魚と考えられる。本流域調査では300 mm以上の個体のみが目視され(後述)、4+歳以上魚であると推定された。本研究では、分流域で捕獲された200mm以下のイトウを幼魚、本流域で確認された300mm以上のイトウを成魚とした。
胃内容物の分析
分流域の調査において、イトウが捕獲された地点で、イトウおよび同所的に捕獲されたイワナとヤマメについてストマックポンプを用いた吐き戻し法により胃内容物を採取し、個体ごとにエタノールで固定した。サンプル数を増やす目的から、調査地点周辺で捕獲された個体も追加した。胃内容物の分析に用いた標本は、イトウ29個体(平均値 ± SD, 尾叉長105.8 ± 20.5 mm, 体重12.9 ± 7.2 g)、イワナ31個体(尾叉長115.6 ± 21.7 mm、体重17.8 ± 10.2 g)、ヤマメ21個体(尾叉長105.6 ± 27 mm、体重16.2 ± 12.5 g)である。調査時にエタノールで固定した胃内容物を実体顕微鏡下で餌生物を目レベルで判別し、個体ごとに分類ごとの餌生物の個体数および乾燥重量を計測した。
統計解析
分流域および本流域の調査において、イトウ生息地点と非生息地点の間に各物理環境の平均値に有意差が認められるかを調べるためマン・ホイットニーのU検定を用いて解析した。また、サケ科魚類の微生息環境に重要と考えられる流速については、他のサケ科魚類を対象として比較し、イトウの選好性との相違についてクラスカル・ウォリス検定を用いて解析した。
分流域および本流域の調査において、各調査地点で計測した物理環境(分流域調査:水温偏差、濁度、川幅、水深、流速、底質粒径、カバー割合;本流域調査:川幅、水深、流速、底質粒径、カバー割合)に関して主成分分析を行った。また、主成分得点をイトウ生息地点と非生息地点で分けt検定を行い、主成分得点の平均値に差があるか検討した。イトウの生息密度と主成分得点の相関関係についてスピアマンの順位相関係数を求めて検討した。
分流域における生息状況と物理環境
分流域調査ではイトウは幼魚のみ、その他魚類はヤマメ、イワナ、フクドジョウBarbatula oreas、スナヤツメLethenteron reissneri、ハナカジカCottus nozawae、ヤチウグイRhynchocypris percnurus sachalinensis、ドジョウ属Misgurnus sp.、ウグイ属Pseudaspius spp.、ウキゴリ属Gymnogobius sp.、ヨシノボリ属Rhinogobius sp.が捕獲された。イトウは23個体が捕獲され、尾叉長69-137mm、体重3.1-26.1gであった。サケ科魚類としては、イワナ183個体(尾叉長61-387 mm、体重2.1-170.3 g)、ヤマメ45個体(尾叉長61-176 mm、体重2.2-65.1 g)が捕獲された(付録1 図2)。調査地点30地点のうちイトウが捕獲されたのは14地点で、生息地点のイトウ生息密度は0.025-0.179個体/m2であった。ヤマメが捕獲された22地点のヤマメ生息密度は0.014-0.197個体/m2、イワナが捕獲された23地点のイワナ生息密度は0.033-1.465個体/m2であった。いずれの種も調査範囲内において偏りなく出現し、上流方向や下流方向に偏った流程分布は示さなかった。
イトウ生息・非生息地点間で物理環境を比較したところ、川幅(P = 0.007)、濁度(P = 0.009)、流速(P < 0.001)、底質粒径(P = 0.017)で有意差が認められ、イトウ幼魚は川幅が狭く、濁度が高く、流速が極めて遅く、底質粒径が小さい場所に生息する傾向がみられた(図2)。イトウ生息・非生息地点の調査時の水温はそれぞれ16.6 ± 3.5℃(平均値 ± SD)および15.5 ± 2.4℃であった。分流域では調査地点によって水温が異なり、本流との水温偏差で-5.1℃から+3.4℃と変動幅が認められたが、イトウ生息の有無と水温の関係は見出されなかった(P = 0.918、付録1 図3)。
分流域の物理環境について主成分分析を行った結果、PC1において生息区と非生息区の間で平均値に有意差が認められた(図3、t = 4.95, df = 28, P < 0.001)。PC1は、流速、川幅、水深、底質粒径の値が大きく、濁度の値が小さい主成分であった。PC1の値が小さい地点でイトウ幼魚の生息密度が高くなる傾向にあり、生息密度とPC1の間には有意な負の相関が認められた(図 4、rs = -0.714, P < 0.001)。PC2以降の主成分ではイトウ幼魚の生息有無との関係性は認められなかった。
他のサケ科魚類が生息していた地点の流速はイトウが生息していた地点よりも有意に速く、ヤマメはもっとも速い流速環境を利用していた(図5、P = 0.002、イトウ:1.6 ± 2.0 cm/s、イワナ:10.1 ± 12.8 cm/s、ヤマメ:16.2 ± 14.8 cm/s)。
本流域における生息状況と物理環境
本流域調査では24個体のイトウが確認され、そのすべての個体が体長300-800 mmの成魚であった(付録1 図4)。イワナおよびヤマメについては、分流調査で捕獲された尾叉長60-200 mmの小型個体も含め多数確認されたが、イトウについては60-200 mmの幼魚は本流域でまったく確認されなかった。なお、潜水調査時には確認できなかったために定量的なデータは得られなかったが、体長50mm弱のイトウ稚魚(0+歳)が、本流域のごく浅い岸際で陸上から時折目視された。調査地点21地点のうちイトウ成魚が確認されたのは11地点で、生息密度は0.004-0.064個体/m2であった。イトウ成魚は調査範囲内において偏りなく出現し、上流方向や下流方向に偏った流程分布は示さなかった。
イトウ成魚の生息・非生息地点で物理環境要素を比較したところ、水深(P < 0.001)、カバー割合(P < 0.001)で有意差が認められ、イトウ成魚は水深が深く、カバー割合が大きい場所に生息する傾向がみられた(図6)。イトウ成魚が確認された地点の流速は平均24.6 m/s(範囲17.1-45.6 m/s)であり、生息・非生息地点のどちらも流れがある地点であった。
本流域の物理環境について主成分分析を行った結果、PC1でイトウ生息区と非生息区の間で、平均値に有意差が認められた(図 7、t検定, t = 2.62, df = 19, P = 0.017)。PC1は、水深、底質粒径、カバー割合の値が大きい主成分であった。PC1の値が大きい地点でイトウ成魚の生息密度が高くなる傾向にあり、生息密度とPC1の間には有意な正の相関が認められた(図8、rs = 0.554, P = 0.009)。PC2以降ではイトウ成魚の生息有無との関係は認められなかった。
幼魚の胃内容物
分流域で捕獲されたイトウ幼魚の胃内容物は、魚類・両生類やスジエビPalaemon paucidensといった大型の水生動物の割合が高く、イワナやヤマメと比較すると、甲虫目Coleoptera、鱗翅目Lepidoptera、ハチ目Hymenopteraおよびカメムシ目Hemipteraなどの陸生落下昆虫の割合が少ない傾向にあった(図9)。大型の餌生物である魚類と両生類には、それぞれヨシノボリ属とエゾサンショウウオ幼生Hynobius retardatusが含まれ、イトウ幼魚の胃内容物から確認された魚類16個体のうち少なくとも12個体がヨシノボリ属であった。一方、小型の動物プランクトンを捕食していたイトウ幼魚も3個体が確認された。イトウ幼魚の餌生物全体に占める動物プランクトンの重量割合は高くないが個体数割合は高く(重量0.4%, 個体数40.1%)、分類群にはミジンコ目Cladoceraとケンミジンコ目Copepodが含まれ、その大半(98%)がミジンコ目であった。
本研究では、網状流路の形態が発達し自然状態が残されている氾濫原に形成された分流(側流)および本流(主要流路)におけるイトウの生息密度調査と物理環境調査により、イトウの幼魚と成魚が夏季に選択的に利用する物理環境の特性を示すことができた。さらに、イトウ幼魚の餌生物を同所的に生息するサケ科魚類と比較を行うことで、イトウ幼魚の生態特性についても新たな知見が得られた。
イトウの幼魚は流速、川幅、水深、底質粒径の値が小さく、濁度が高いという、本流的な環境からかけ離れた地点に生息することが明らかとなった(写真、付録1 図5)。イトウ幼魚は他のサケ科魚類が好む流れのある地点ではなく、氾濫原に形成される止水的環境を選択的に利用していることを示していると考えられる。とくに流速に関しては0-3.6 cm/s程度の極めて流れの緩やかな環境を利用していることがわかった。このようにイトウは幼魚期に氾濫原に形成される水路を利用していたことから、土地や農地開発等の影響により氾濫原環境が減少することは、イトウの個体数ひいては個体群の減少につながることが示唆される。
江戸・東(2002)および佐川ほか(2003)はイトウ幼魚が生息していない環境との比較をしていないが、イトウ幼魚が生息していた環境の特性として、流速が極めて遅い環境であると報告しており、本研究の結果と一致する。江戸・東(2002)によると、産卵床の形成された本流において夏に浮上したイトウの稚魚は、秋頃になると氾濫原に多くみられる直接本流に注いでいる小水路に入り込むと述べており、本研究でイトウ幼魚が生息した分流域と一致する。ブトカマベツ川における観察でも、秋には尾叉長40-70 mmに成長した稚魚(0+歳)が分流に入り込む様子が確認されている(宇野ほか 未発表)。本研究において、イトウ幼魚は分流域であっても流速が3.6m/s以上の環境では全く生息が確認されなかったことから、流速が極めて遅い環境に対して強い選択性をもつと考えられる。
一方、イトウの成魚は流れのある本流域で確認され、水深、底質粒径、カバー割合の値が大きいという、隠れ家の多い深い淵を利用していると考えられる(写真、付録1 図5)。イトウ成魚は夏季生息域として樹冠被覆度が高く、流速が遅く、水底面積が大きい淵を選択的に利用することが報告されており(佐川ほか 2002)、本研究でも同様の傾向がみられた。イトウ成魚は氾濫原に生息しているとはいえないが、稚魚、幼魚等未成魚の生息、成長に氾濫原環境は欠かすことはできず、また成魚も護岸工事等、河川の直線化の影響により、流速が遅くカバーがあり、大きい淵である環境が減少することで負の影響を受けている可能性がある。多くのサケ科魚類において、倒木などのカバーが多い淵が重要な生息場所となることが指摘されている(永山ほか 2008)。例えば、北海道北部の増幌川や北海道東部の標津川において、水深が深く、倒木などのカバーが多い淵がサクラマスの重要な生息場所となっていると報告されている(Edo and Suzuki 2003; Nagayama et al. 2009)。本研究で明らかとなったブトカマベツ川のイトウ成魚の生息環境は、これまでサケ科魚類の成魚にとって重要と指摘されてきた環境特性と一致する。
本研究では尾叉長200 mm前後のイトウを捕獲、目視することができなかった。イトウは孵化後、氾濫原を利用しながら成長し、下流方向へ分散していく可能性が指摘されている(佐川 2006)。そのため、本研究で確認できなかった尾叉長200 mm前後の個体は、調査範囲外であったブトカマベツ川の下流域やその下流に位置する朱鞠内湖に生息している可能性も考えられる。本河川に生息しているヤマメは、1+または2+歳でスモルト化し朱鞠内湖を索餌回遊する湖沼回遊型となり、産卵のために河川に遡上することが知られている(Tamate and Maekawa 2000)。また、ブトカマベツ川本流に設置された下り簗により、尾叉長15 cm程度のスモルト化したイトウが春季に捕獲されており(前川光司 私信)、実際、朱鞠内湖では成長したイトウが釣りの対象となっている。
イトウ幼魚の胃内容物はイワナやヤマメと比較して、陸生落下昆虫の割合が少なく、魚類・両生類やスジエビなどの甲殻類といった大型の動物や動物プランクトンも捕食していることが明らかとなった。イワナやヤマメなどの河川性サケ科魚類は、基本的には上流に向かって流れの中で定位し、川の流れによって運ばれてくる水生昆虫や陸生昆虫などを採餌する待ち伏せ型であることが知られているが(中野・谷口 1996; 中野 2003)、流れのほとんどない環境に生息するイトウ幼魚は、大型の動物や底生動物、動物プランクトンなどを探索的に捕食しているのではないかと推察される。定位行動を示す一般的なサケ科魚類には採餌をめぐる干渉型競争がみられ、同種・異種個体間で順位やなわばりが形成されるが(中野・谷口 1996; 中野 2003)、久保(1990)によるイトウの水槽飼育での観察によると、イワナやニジマスOncorhynchus mykissとは対照的にイトウの幼魚は競争や闘争の行動は示さず、他のサケ科魚類に見られるようななわばり性を示さないという。さらに、イワナとイトウの幼魚を同じ水槽で飼育すると、イトウはイワナによる攻撃を受けて次々と死亡に至ったと述べられている。イトウは、同所的に生息する他のサケ科魚類と異なる採餌戦略をとった結果、異なる餌生物と流速環境を利用することにより他のサケ科魚類との共存を可能にしていると考えられる(佐川ほか 2003; 鈴木 2016)。
イトウは肉食であり、強く大きな両顎と鋭利な歯を多くもち、沿岸域では大型の魚類(Edo et al. 2005; 鈴木 2016)、河川では魚類や小型哺乳類等を捕食することが知られている(木村1966; 中野 1992; Ohdachi and Seo 2004)。佐川ほか(2003)は、北海道北部の河川で採集されたヤマメの胃内容物には陸生落下昆虫が多く含まれるのに対し、イトウ幼魚はカゲロウ目や魚類(フクドジョウ、エゾトミヨPungitius tymensis、イトウ稚魚)を捕食していたと報告している。木村(1966)も北海道東部のイトウの食性を分析し、全長14-15 cmから魚類を捕食するようになると述べている。下田ほか(2020)は本研究を行った朱鞠内湖流入河川において得られたイトウ1+歳幼魚の窒素安定同位体の分析によって、魚類を捕食していた可能性を指摘している。本研究でも、小さい個体では尾叉長79 mmから胃内容物に魚類が含まれており、イトウは幼魚期から魚食性を示した。ブトカマベツ川では、夏季に朱鞠内湖から遡上してくるヨシノボリ属がイトウ幼魚にとって重要な餌資源と考えられた。なお、浮上直後のイトウ稚魚(尾叉長31-33 mm, n = 10)は主にユスリカの幼虫・成体を利用していた(遠藤・森田 未発表)。
本研究では動物プランクトンがイトウ幼魚(3個体)、イワナ(1個体)およびヤマメ(1個体)の胃内容物から出現した。これまでプランクトンがイトウ幼魚の胃内容物から報告されたことはなく、著者らが知る限り初の報告である。また、淡水域のサケ科魚類においても、湖に生息するイワナ属魚類などの例を除くと(e.g., Malmquist et al. 1992)、河川に生息するサケ科魚類において胃内容物から動物プランクトンが出現した例はほとんど見当たらない。氾濫原の止水域ではプランクトンが豊富な環境も形成されるが(Uno et al. 2022)、そうした環境下ではサケ科魚類はプランクトンも利用することが明らかとなった。また、ブトカマベツ川流域におけるプランクトン相としては、ケンミジンコ目Copepodが90%以上であるの対し(Uno et al. 2022)、イトウが捕食していたプランクトンは98%がミジンコ目Cladoceraと著しく偏っていた。コイ科魚類においても同様のミジンコ目に対する選択性が知られており、ケンミジンコ目の高い遊泳速度が魚類の捕食率の低下と関係しているのかもしれない(Hambright and Hall 1992)。
本研究の調査ではイトウ幼魚は氾濫原に形成される流速が極めて遅い環境を選択的に利用するのに対し、イトウ成魚は流れのある本流で深くカバーのある環境を選択的に利用していた。このようにイトウは成長段階に応じて氾濫原に形成される様々な水域を横断的に利用していることが明らかとなった。回遊性のサケ科魚類の保全には、これまで縦断方向の連続性に注目されることが多かった(森田・山本 2004; 福島 2005)。しかし、イトウの個体群存続性を高めるためには、幼魚期を含めた生活史段階を考慮して横断方向の連続性の保全にも着目し、氾濫原環境を減少させるような川の直線化工事や護岸工事の手法を見直す必要がある(石山ほか 2017)。
謝 辞
北海道大学雨龍研究林の中路達郎林長、坂井励氏、原悠子氏をはじめとするスタッフの皆様には本研究を実施するにあたり多大なるご協力いただきました。朱鞠内湖淡水漁業協同組合の中野信之氏にはさまざまな便宜を図っていただいた。元北海道大学雨龍地方演習林の前川光司先生からは当時のイトウに関する貴重な情報を頂いた。2名の匿名の査読者には原稿の改善に資する有益な助言を頂いた。ここに感謝の意を表します。本研究はJSPS科研費 22K14915、22H02694の助成を受けたものです。
著者情報
ORCID
Hiromi Uno https://orcid.org/0000-0002-9343-3061
Osamu Kishida https://orcid.org/0000-0002-2663-8155
Kentaro Morita https://orcid.org/0000-0002-7803-2438
引用文献

北海道北部朱鞠内湖流入河川ブトカマベツ川と各調査地点の位置。
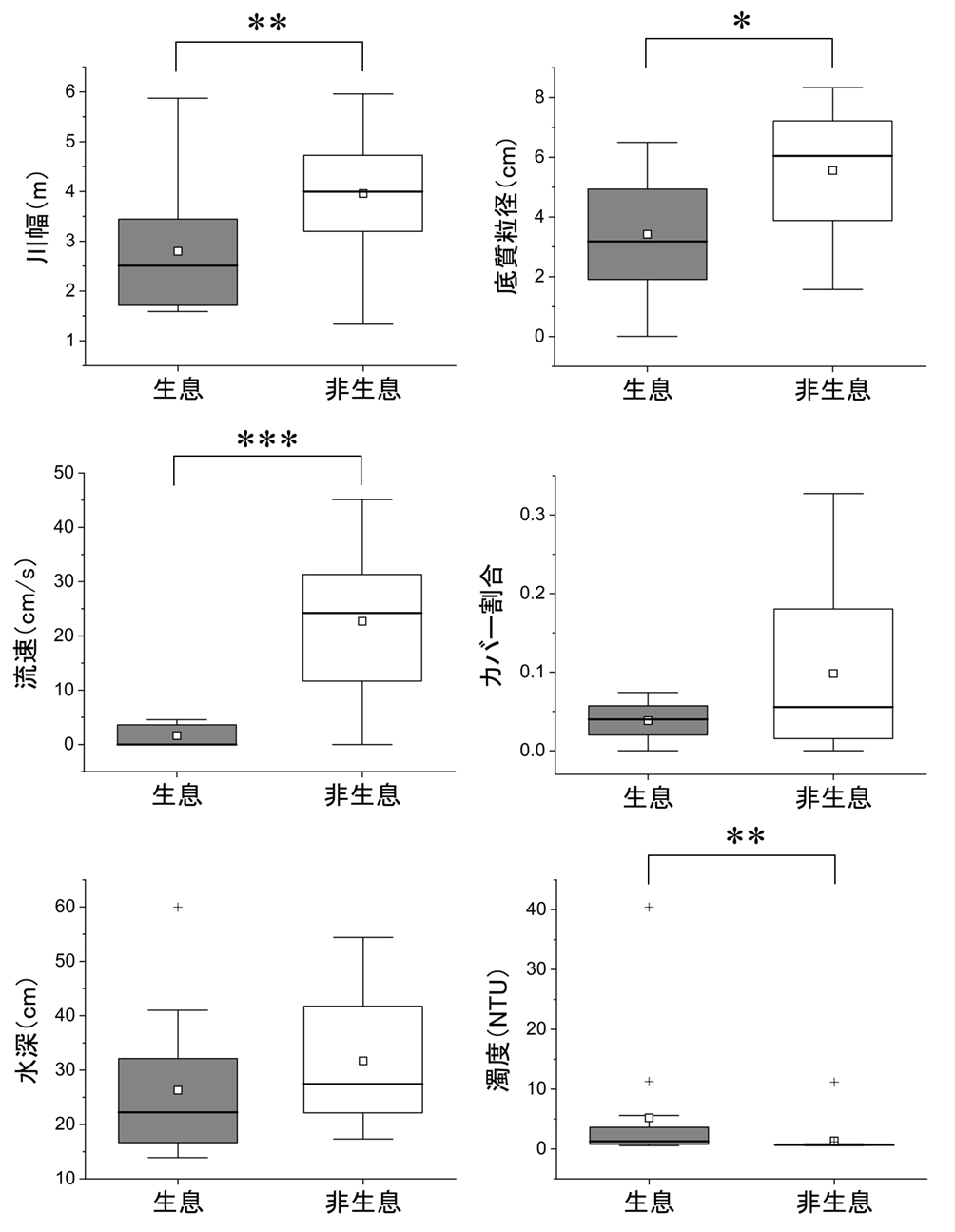
分流域調査におけるイトウ生息・非生息環境の比較。太線は中央値、白四角は平均値、箱の上端と下端はそれぞれ第1四分位と第3四分位、エラーバーは範囲を示す。第1四分位と第3四分位の間の1.5倍を超える範囲は外れ値として+記号で示す。*P < 0.05; **P < 0.01; ***P < 0.001。
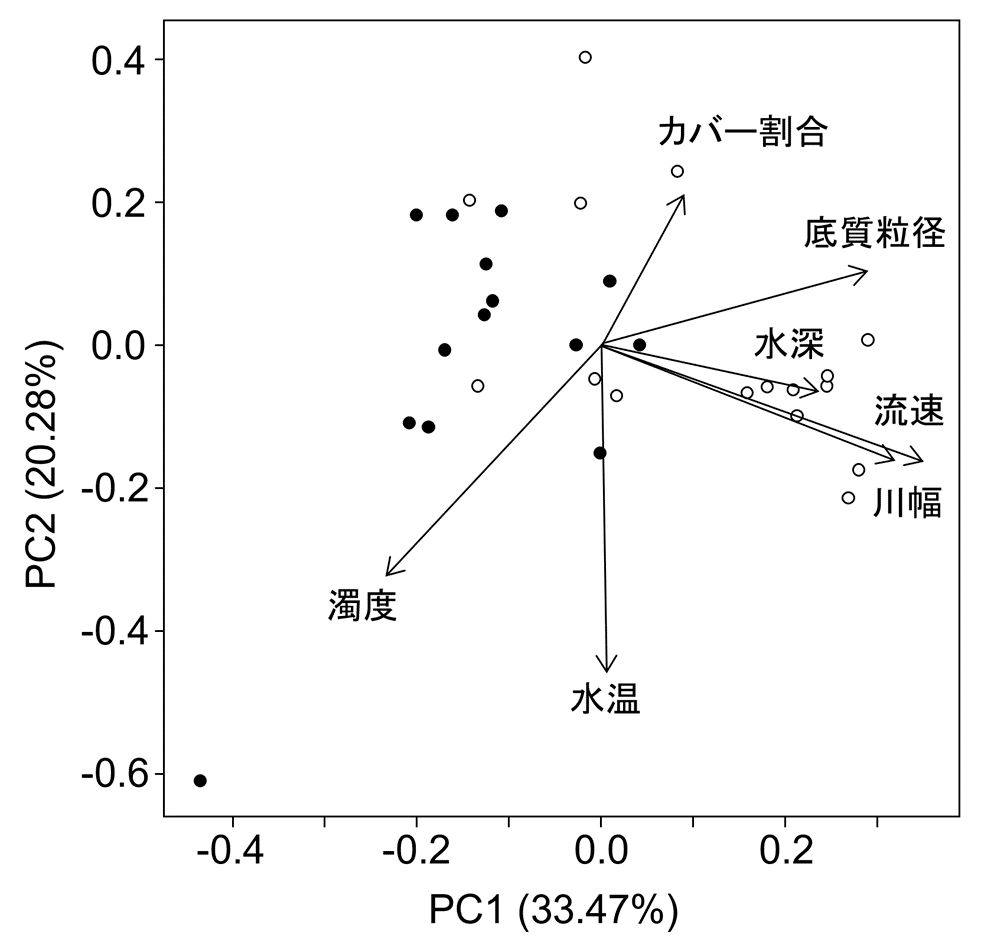
分流域調査における物理環境の主成分分析の結果。黒丸:イトウ幼魚生息地点、白丸:イトウ幼魚非生息地点。

分流域物理環境のPC1の主成分得点とイトウ幼魚生息密度の関係 黒丸:イトウ幼魚生息地点、白丸:イトウ幼魚非生息地点。
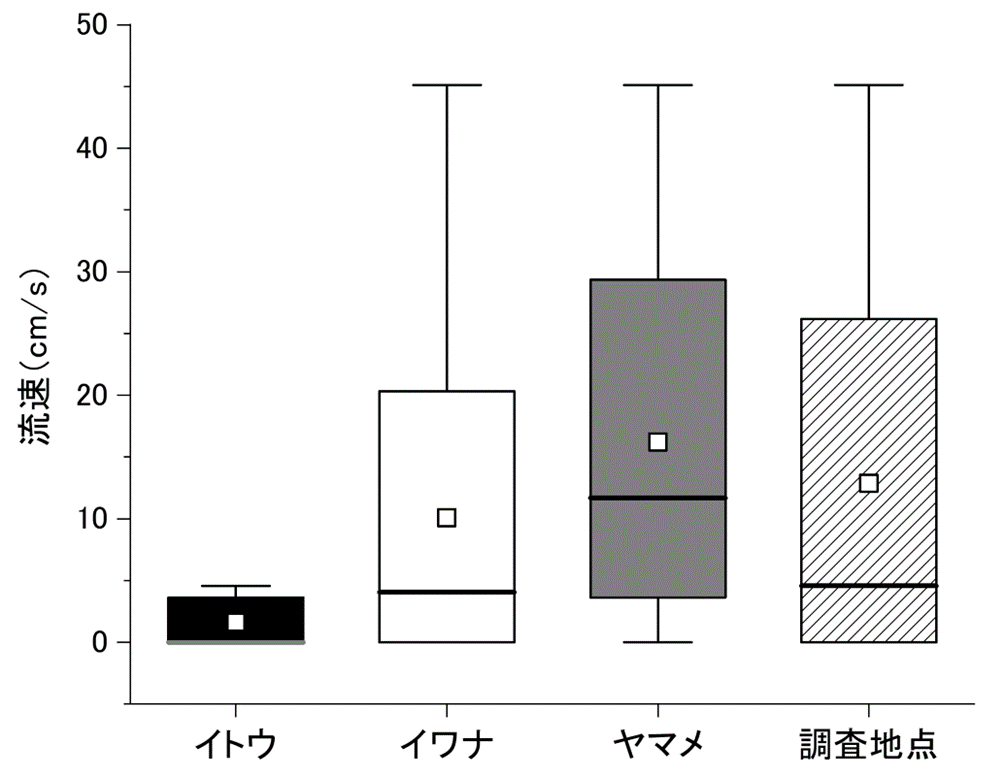
分流域調査におけるイトウ、イワナ、ヤマメ生息地点および全調査地点の流速分布。図の見方は図2に同じ。
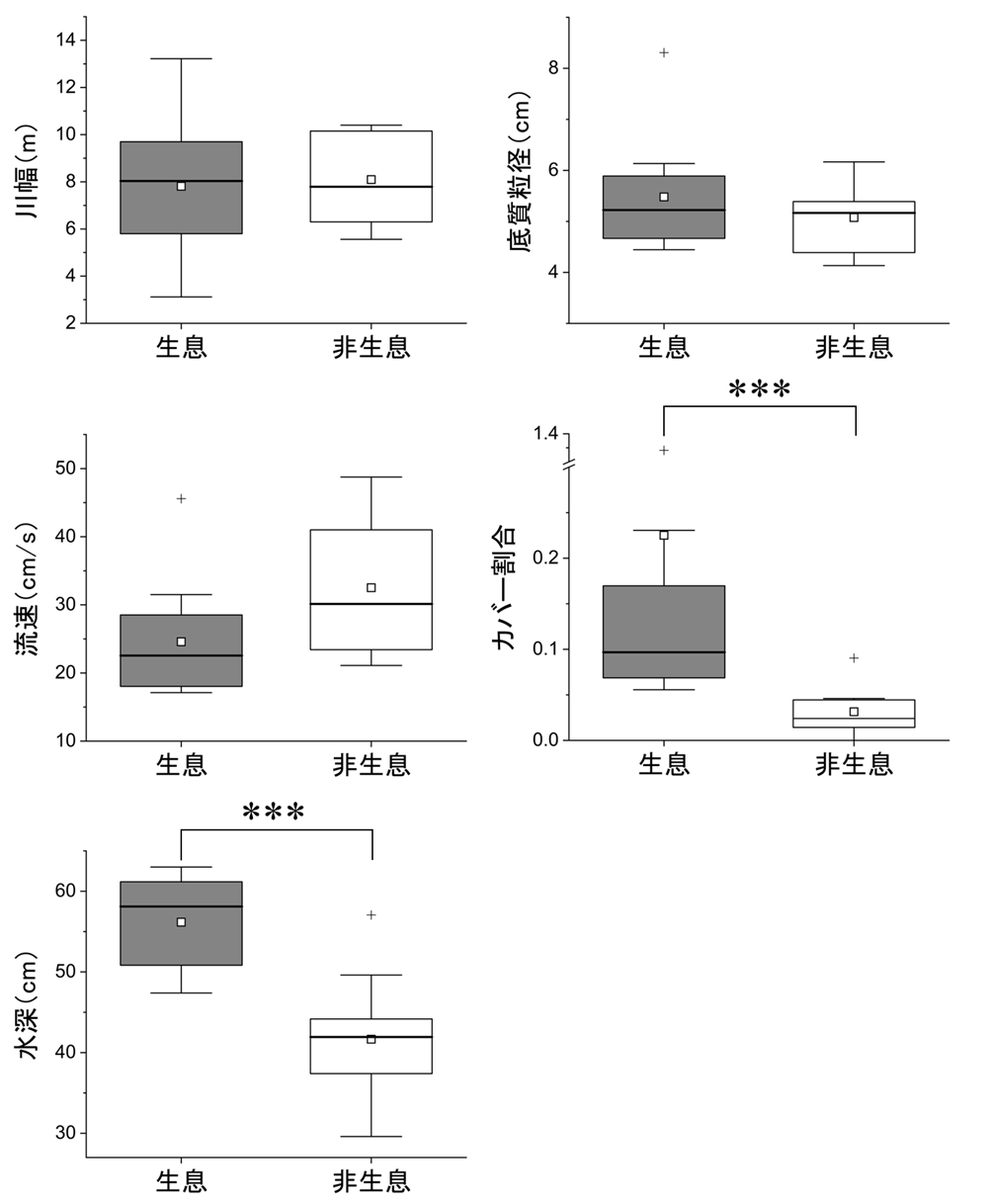
本流域調査におけるイトウ生息・非生息環境の比較。図の見方は図2に同じ。
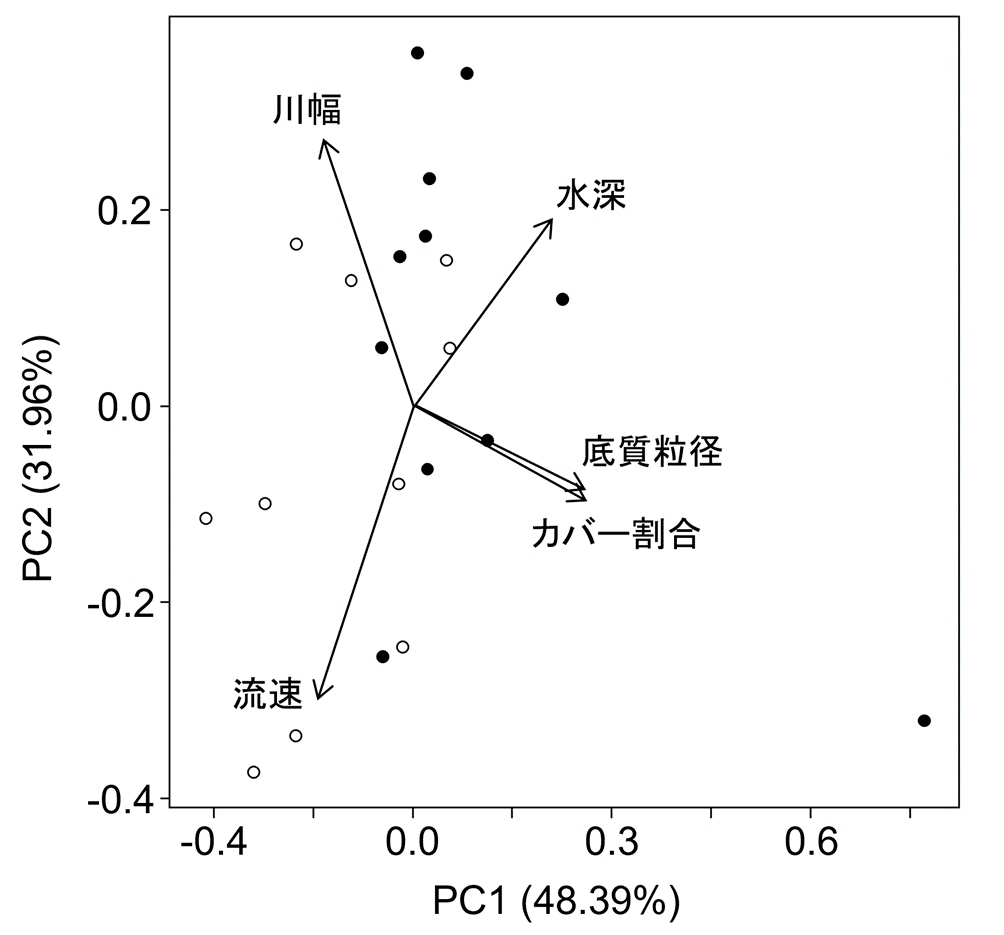
本流域における物理環境の主成分分析の結果。黒丸:イトウ成魚生息地点、白丸:イトウ成魚非生息地点。
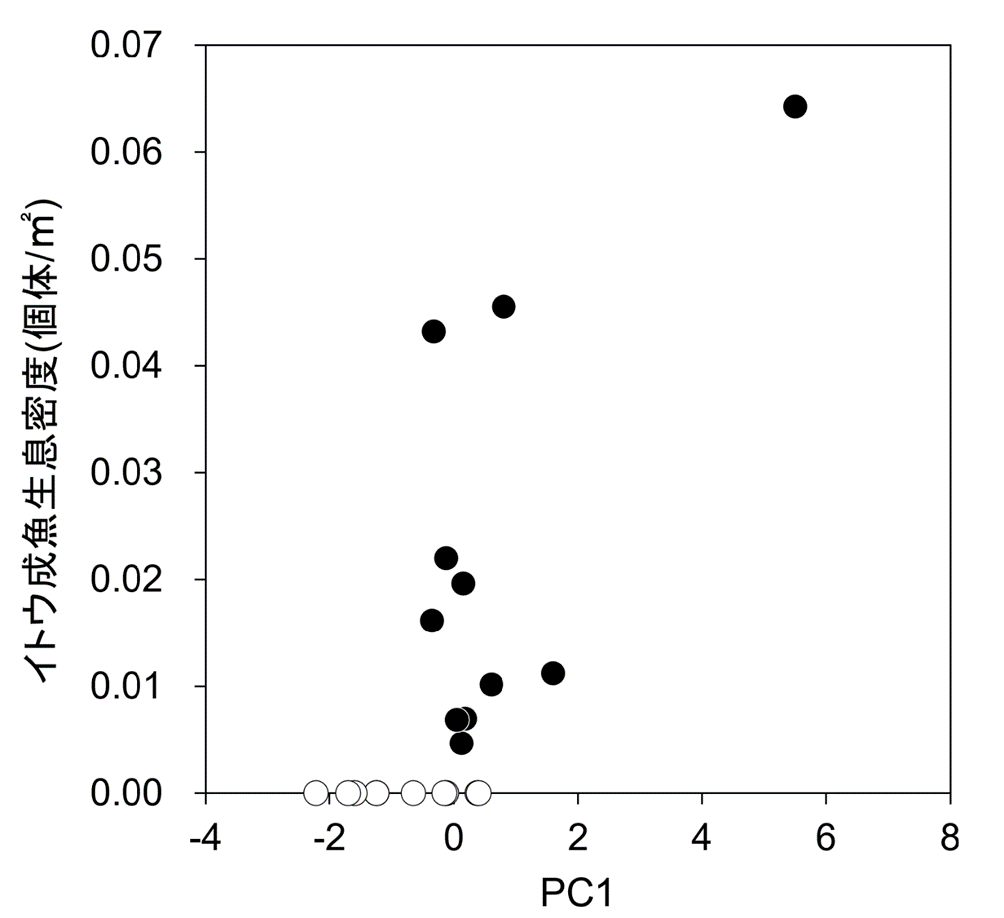
本流域物理環境のPC1の主成分得点とイトウ成魚生息密度の関係。黒丸:イトウ成魚生息地点、白丸:イトウ成魚非生息地点。

図9
イトウ幼魚、イワナおよびヤマメの個体別の分類群毎の胃内容物重量。縦軸は各個体の尾叉長を示す。

