Processing and Products
Influence of Long-Distance Transportation Under Various Crating Densities on Broiler Meat Quality During Hot and Humid Weather
2020 年 57 巻 3 号 p. 246-252

詳細
2020 年 57 巻 3 号 p. 246-252
The objective of the study was to investigate the effect of transportation distance and crating density on broiler meat quality during hot and humid weather. Breast meat samples were collected from broilers (Ross-308) after transportation for 80, 160, and 240 km at crating densities of 10, 12, and 15 birds per crate. The broilers were transported in plastic crates of 0.91 × 0.55 × 0.31 m (length × width × height) in summer at an ambient temperature of 27.2–33.6°C and relative humidity of 52.7–62.9%. The birds were then slaughtered, and the meats' physicochemical characteristics, marination and cooking attributes were measured. Meat pH at 2 hours (h) postmortem (PM) was significantly higher in birds transported for 80 and 160 km compared to those transported for up to 240 km, while meat pH at 24 h PM was higher in birds transported for up to 160 km compared to the other treatments; however, no effect of different crating densities on meat pH and drip loss was observed. Meat losses such as drip loss, thaw loss, cooking loss in raw and marinated breast, and meat shear force were significantly higher in birds transported for up to 240 km compared to the other treatments. An increase in crating density above 12 birds per crate also increased thaw loss, whereas crating density of 12 birds per crate and higher increased cooking loss and meat shear force. Meat lightness also increased significantly with the increase in transportation distance above 160 km and crating density above 12 birds per crate. In conclusion, transportation of broilers for more than 80 km with crating density higher than 12 birds per crate during summer was associated with severe losses in yield and deterioration in physical appearance and functional characteristics of meat.
Global warming is severely affecting the climatic conditions particularly of developing countries such as Pakistan by widening the gap between winter and summer temperatures (Abbas, 2013). High ambient temperature coupled with high humidity not only reduces growth performance but also threatens the well-being of broilers during their journey towards abattoirs (Aldridge et al., 2019). Temperature and humidity variations during transportation can cause high mortalities (Arikan et al., 2017), oxidative damage to muscles (Estévez, 2015), and can also reduce physical appearance and functional characteristics of meat by inducing protein denaturation, pale color, and drip loss (Wilhelm and Maganhini, 2010; Xing et al., 2015). The extent of deterioration during transportation depends on the microclimate of the vehicle compared to the direct effect of the ambient environment (Simões et al., 2009).
Broiler transportation trucks usually depend on passive air movements to dissipate the accumulated heat and moisture (Eigenberg et al., 2009). Non-uniform movement of air in these open-sided trucks has been associated with the development of a thermal core, in which some regions of the truck, particularly the middle and rear ones, have a higher temperature than other regions (Simões et al., 2009). Length of transportation interacts with crating density to determine the microclimate of the vehicle and the level of broiler stress under different climates (Bedanova et al., 2006; Queiroz et al., 2015; Nikbin et al., 2016). The negative effect of an increase in transportation distance on meat quality has already been reported in various geographical ranges and climatic zones (Bianchi et al., 2006; Yalçin and Güler, 2012; Aral et al., 2014). However, some scientists have reported contradicting findings; Zhang et al. (2009), for example, reported that broiler meat quality was not affected by transportation distance.
Ambient temperature during transport is one of the critical factors in determining optimal crating densities. However, the effect of broiler crating densities, particularly in hot climates, are among the least studied subjects. A study conducted regarding this aspect reported a significant increase in stress parameters at higher crating densities based on blood biochemical profile and physiological implications (Suchy et al., 2007). Delezie et al. (2007) showed that the increase in crating density (0.0350 m2/broiler and 0.0575 m2/broiler) resulted in higher body temperature and blood corticosterone level without any significant effect on meat quality of broilers in Belgium. Moreover, an excessively low crating density can affect transportation economics and increase injuries, whereas an excessively high density can lead to hyperthermia and other quality deteriorations. Therefore, the evaluation of crating density under different temperatures is a critical aspect that must be investigated. The present study was conducted to determine the effect of different transportation distances and crating densities on meat quality during hot and humid summer.
The experiment was conducted at the Department of Poultry Production, University of Veterinary and Animal Sciences, Lahore, Pakistan. The broilers were transported in the vicinity of District Kasur (Pattoki City) and District Lahore, Punjab, Pakistan in laps of 40 km.
Experimental BirdsIn this experiment, ROSS-308 broilers (straight-run; body weight 1900–2050 g; age 35 days) were selected from a commercial broiler farm after applying a four hour (h) long feed withdrawal before catching. During the growing phase, the broilers were fed with commercially available broiler ration (strain specific) and raised on litter floor in an environmentally controlled broiler house under optimum management conditions. Broiler samplings were based on three transportation distances (i.e., 80 km, 160 km, and 240 km) with three crating densities {Low= 10 birds (0.05 m2/bird), Medium=12 birds (0.042 m2/bird), and High= 15 birds (0.033 m2/bird) per crate}. The birds were transported in commercial trucks carrying different crating densities in loose plastic crates (Engi Plastic Industries Pvt. Ltd., Sundar, Pakistan) with dimensions of 0.91×0.55×0.31 m (length×width×height) at an average speed of 45–50 km/h during late night to early morning. To reduce the effect of truck microenvironment, the samples were collected from random locations of the trucks. All the procedures and bird handling protocols were used according to guidelines and standards after approval from the Ethical Review Committee of University of Veterinary and Animal Sciences, Lahore (Approval No. DR/916-2017).
Climatic IndicesThe experiment was conducted during summer (Mid-August). Temperature and relative humidity were recorded with a digital weather tracker (Kestrel 4500® NV, Nielsen-Kellerman, USA). The temperature range and relative humidity measured at various times and distances remained between 27.2–33.6°C and 52.7–62.9%, respectively. The birds were transported without any misting or water sprinkling during the whole journey.
Parameters StudiedAt the end of transportation, the crates were unloaded manually and three birds from each replicate were selected (10 replicates per treatment), slaughtered with a knife, defeathered, and eviscerated. After recording some initial parameters, carcasses from each treatment were placed in chilled water to reduce the temperature, and the following parameters were then recorded.
Meat pHThe pH of individual breasts was measured from three locations using an electronic pH meter (ProfiLine pH 3110 SET 2, WTW, Germany), as measured by López et al. (2011). The pH was measured from the warm carcasses at 15 minutes Post Slaughter (PS), and from deboned breast samples stored at 4°C in a display chiller at 2 h and 24 h PS (S80100VVC, Tecnodom S.P.A., Italy).
Meat ColorThe breast meat (pectoralis major) was separated from each carcass, placed in polystyrene food grade trays covered with cling film (250 mm thickness), and kept in a display chiller (S80100VVC, Tecnodom S.P.A., Italy) at 4°C. Meat color (L*=lightness, a*=redness, b*=yellowness, c=chorma, h=hue) was measured at 24 h postmortem (PM) employing the Commission International de l'Eclairage (CIE, 1976) L*a*b* system using a chroma meter (Minolta® CR-410, Konica Minolta, Inc. Japan).
Drip and Thaw Loss (%)Drip loss (%) was measured by hanging each breast muscle (weight 135–150 g) by a thread in a polythene bag under atmospheric pressure for 24 h at 4°C, as described by Honikel et al. (1998). Drip loss (%) was calculated as the ratio of the difference between the initial and final weight of a sample to the initial weight of the sample. Thaw loss (%) was measured following the procedure of Dadgar et al. (2010), in which the frozen breast (slow freezing) was kept for thawing at 4°C overnight. Thaw loss (%) was calculated as the ratio of the difference between the samples' weight before freezing and after thawing and blotting to weight before freezing.
MarinationMarination of six breast fillets (uniform weight) per treatment (kept at 4°C) was performed with 4% brine solution (chilled overnight) using the methodology of Yusop et al. (2010) with slight modifications. Breast samples were immersed {i.e., batch of 10 pre-weighed (w1) fillet} in marinade solution and manually tumbled for 15 minutes, providing equal time to all treatments, for maximum possible absorption of brine. After performing marination, the fillets were reweighed (w2) to determine marinade uptake. The fillets of each treatment were placed in polystyrene trays and held overnight at 4°C. Then, individual fillets were reweighed (w3) to calculate marinade retention using the formulas:
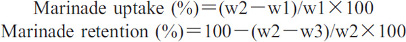 |
For measurement of cooking losses, the method described by Honikel (1998) was used with slight modifications. The breast samples (raw and marinated) were individually packed in labelled polythene zipper bags (food grade). The bags with the weighed samples (w3) were placed in a water bath (WNB45 Memmert GmbH, Schwabach, Germany) operating at 80°C. The samples were then cooked until the core temperature of the thickest part reached 72°C. Temperature was monitored with a digital thermometer (TP-101, China). After cooking, the fillets were cooled, blotted dry, and reweighed (w4) to calculate the losses during cooking using the formula:
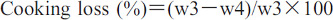 |
The Warner Bratzler Shear Force Value (N/cm2) of raw cooked and marinated cooked meat was analyzed using Texture analyzer (TA. XT plus® texture analyzer, Stable Micro System, UK). Three rectangular blocks, each with a cross-section of 1 × 1 cm2 and fiber direction parallel to a long dimension of 2–3 cm, were cut from each cooked fillet (Dadgar et al., 2010). Each rectangular strip (2–3 cm long) was cut 3–4 times and mean value was calculated. The fibers were cut with a V slot blade at a cross speed of 2 mm/s during shear.
Statistical AnalysisThe data were analyzed using the two-way analysis of variance technique with a completely randomized design under factorial arrangement. Crating densities and transportation distances were used as main effects in this experiment. Tukey's HSD test (Tukey, 1953) was used in the statistical package SAS 9.4 (SAS Institute, 2013) for the separation of significant means at p<0.05.
Initial meat pH (i.e., at 15 minute PS) was non-significantly different among different transportation distances (p=0.1455), crating densities (p=0.3290), and their interactions (p=0.9851). At 2h PM, pH in birds subjected to 240km transportation was lower (p<0.0001) than in those transported for 80 and 160 km (Table 1). Similarly, pH at 24 h PM (ultimate pH) was significantly higher in birds subjected to 160 km transportation, followed by those transported for up to 80 km, and then by those subjected to 240 km transportation (Table 1). A comparable (p>0.05) initial pH was observed in all treatment groups, but thereafter a rapid decline in pH (difference between initial and 2 h PM pH) was observed in birds subjected to longer transportation (i.e., 240 km), whereas a normal decline occurred in the other treatments. This finding could be attributed to the birds' inability of coping with the stress produced due to the accumulation of heat during the long distance transportation at higher ambient temperature, which resulted in an accelerated pH fall along with an increase in the extent of pH fall (ultimate pH) during rigor mortis development (Dadgar et al., 2010; Jiang et al., 2015). Zhang et al. (2012) explained that low ultimate pH during heat stress results from the high production of lactic acid in the muscles. Under stress, the autonomous sympathetic nervous system releases catecholamines, which trigger the “fight and flight” response in birds and stimulate voltage-gated calcium channels to release high amounts of calcium into the sarcoplasm (Spinosa, 2006). The sudden increase of Ca2+ ions in the sarcoplasm then increases the speed of postmortem utilization of muscle adenosine triphosphate (ATP) and glycogenolysis, resulting in accelerated rates of decline in muscle pH (Rübensam, 2000). Similar to our findings, Henckel et al. (2000) also reported that the rate of pH decline in stressed animals can be two to four (or sometimes almost six) times quicker than in non-stressed animals, as observed in 240 km transported birds at 2 h PM in the present study (Table 1). Not only heat stress but also muscle glycogen contents are critical in determining meat quality. Some studies have associated the increase in transportation distance with depletion of muscle glycogen and low lactic acid accumulation, which results in higher than normal ultimate pH (Zhang et al., 2009); this is similar to our findings regarding birds transported for up to 160 km compared to those transported for 80 km. Accordingly, a significant decrease in ultimate meat pH with the increase in transportation distance of birds subjected to 30 to 180 km transportation has also been previously reported (Akşit et al., 2006; Oba et al., 2009). In the present study, pH at 2 and 24 h PM showed non-significant differences (p>0.05) among different crating densities and different transportation distance × crating density interactions (Table 1). Similarly, Delezie et al. (2007) also reported no difference between meat pH at two crating densities. This contradiction could be attributed to factors like relatively lower transportation time (1.5 h) and rest after transportation. However, there is lack of published research regarding crating density.
| Parameters | Distance | Crating density | P value | ||||||
|---|---|---|---|---|---|---|---|---|---|
| 80 km | 160 km | 240 km | Low | Medium | High | Distance | Density | Interaction | |
| pH 15 min | 6.57±0.01 | 6.56±0.01 | 6.54±0.01 | 6.57±0.01 | 6.56±0.01 | 6.55±0.01 | 0.1455 | 0.3290 | 0.9851 |
| pH 2h | 6.13±0.03a | 6.11±0.01a | 5.99±0.02b | 6.11±0.03 | 6.06±0.02 | 6.05±0.02 | <0.0001 | 0.0974 | 0.9693 |
| pH 24h | 5.81±0.01b | 5.87±0.01a | 5.77±0.01c | 5.81±0.01 | 5.82±0.02 | 5.80±0.01 | <0.0001 | 0.5146 | 0.8503 |
| Drip loss % | 2.84±0.12b | 2.83±0.10b | 3.46±0.23a | 2.82±0.15 | 3.13±0.17 | 3.19±0.19 | 0.0019 | 0.1275 | 0.0012 |
| Thaw loss % | 12.14±0.43b | 11.09±0.48b | 13.72±0.68a | 12.41±0.49ab | 11.34±0.50b | 13.20±0.70a | 0.0007 | 0.0196 | 0.0018 |
| Raw Breast | |||||||||
| CL % | 22.77±0.77ab | 21.92±0.57b | 23.92±0.65a | 21.23±0.44b | 23.38±0.47a | 24.01±0.89a | 0.0730 | 0.0056 | 0.2065 |
| SF (N/cm2) | 14.06±0.60b | 15.07±0.79b | 18.06±0.52a | 13.52±0.65b | 16.58±0.78a | 17.09±0.53a | <0.0001 | <0.0001 | 0.0002 |
| Marinated breast | |||||||||
| MU % | 15.06±0.89 | 15.16±0.86 | 17.25±0.94 | 14.33±0.70 | 16.55±0.79 | 16.59±1.14 | 0.1316 | 0.1090 | 0.1294 |
| MR % | 95.42±0.40a | 95.11±0.34a | 93.53±0.37b | 95.23±0.33 | 94.64±0.30 | 94.19±0.55 | 0.0010 | 0.1294 | 0.1883 |
| CL % | 24.85±0.25c | 26.14±0.36b | 29.16±0.42a | 25.80±0.45b | 26.61±0.57b | 27.73±0.55a | <0.0001 | 0.0001 | 0.2558 |
| SF (N/cm2) | 6.16±0.26c | 8.59±0.29b | 11.46±0.69a | 8.75±0.60 | 9.44±0.65 | 8.01±0.50 | <0.0001 | 0.0804 | 0.2041 |
Values are least square mean±standard error. Means in a row with no common superscript (a, b, and c) differ significantly at p≤0.05
Cratingdensity: Low=10 birds/crate (0.050 m2/bird), 12 birds/crate (0.042 m2/bird), and 15 birds/crate (0.033 m2/bird)
CL=CookingLoss, SF=Shear Force, MU=Marinade uptake, MR=Marinade retention
Marinated: Breast marinated with 4% brine solution for 15 minutes
In this experiment, drip loss (%), thaw loss (%), and cooking loss (%) in raw and marinated breasts were significantly higher (p<0.05) in birds transported for 240 km compared to the other treatments (80 and 160 km). Whereas, among different crating densities, non-significant differences were observed for drip loss (%), but thaw loss (%) and cooking loss (%) were significantly higher in birds crated at higher density (i.e., 15 birds per crate) followed by those transported at low crating density (i.e., 10 birds per crate), and then by those transported at medium crating density (Table 1). Moreover, transportation distance × crating density interactions were significantly different for drip loss and thaw loss % (Table 2). Results regarding interactions showed significantly higher drip loss (%) in birds transported for up to 240 km at medium and high crating densities compared to all other treatments. Similarly, the interactions also showed that thaw loss (%) increased with the increase in transportation distance and crating density (Table 2). Water is an integral component of muscle cells, captured by proteins in muscle fibers (Bowker, 2017). Previous studies have reported that the amount of water loss from meat and water binding capacity of proteins are dependent on their isoelectric point (Alvarado and McKee, 2007). The isoelectric point is a pH at which there will be no net charge on proteins, and thus they will release the bound water molecules. Actin and myosin are two of the major muscle proteins, and they have isoelectric points between 5.1–5.4 (Bowker, 2017). The farther the muscle pH from the isoelectric point, the higher the charge on the proteins and the water binding capacity, and more intracellular spaces for water will exist (Huff-Lonergan and Lonergan, 2005). Similarly, in the present study, treatments with lower ultimate pH showed higher drip losses (Table 1). Caldara et al. (2012) reported a significant increase in cooking losses when the ultimate pH decreased below 5.9. Similarly, studies in other counties have also reported an increase in drip loss and cooking loss under high ambient temperatures, chronic heat stress (Lu et al., 2010; Wang et al., 2017), and after merely 0.5 h of transportation (Xing et al., 2015). Contrary to our results, a study conducted in Belgium reported no effect of crating density (0.0350 m2/bird or 0.0575 m2/bird) on drip loss (Delezie et al., 2007). This contradiction could be a result of the relatively higher stress level in the present study resulting from the longer transportation distance (80–240 km) under high ambient temperature compared to the 1.5 h transportation period of Delezie et al. (2007). This finding suggests that ambient conditions and transportation distance have major impacts on drip and cooking losses. Regarding raw meat cooking, birds transported for 160 km had higher ultimate pH (Table 1) compared to those transported for 80 and 240 km (Table 1), either due to the higher depletion of glycogen relative to the 80 km of travelling or due to the relatively lower heat exposure relative to the 240 km treatment; therefore, meat, having a high pH, will lose less water, which results in lower cooking losses (Gregory, 2010; Caldara et al., 2012).
| Transportation Distance | Crating density | Drip loss (%) |
Thaw loss (%) |
Shear Force (N/cm2) Raw breast |
|---|---|---|---|---|
| 80km | Low | 3.13±0.32b | 13.86±0.55b | 13.40±1.26c |
| Medium | 2.79±0.10b | 11.55±0.79bcd | 12.20±0.68c | |
| High | 2.60±0.09b | 11.01±0.14cd | 16.58±0.58b | |
| 160km | Low | 2.77±0.25b | 11.17±1.03cd | 11.00±0.39c |
| Medium | 2.74±0.11b | 9.79±0.44d | 17.98±1.27ab | |
| High | 2.98±0.15b | 12.32±0.66bc | 16.21±1.17b | |
| 240km | Low | 2.56±0.21b | 12.21±0.62bcd | 16.16±0.90b |
| Medium | 3.84±0.33a | 12.68±0.96bc | 19.55±0.70a | |
| High | 3.98±0.35a | 16.25±1.23a | 18.48±0.79ab |
Values are least square mean±standard error. Means in a column with no common superscript (a, b, c, and d) differ significantly at p≤0.05
Cratingdensity: Low=10 birds/crate (0.050 m2/bird), 12 birds/crate (0.042 m2/bird), and 15 birds/crate (0.033 m2/bird)
Marinade uptake (%) was not affected by different transportation distances (p=0.1316), crating densities (p=0.1090), and their interactions (p=0.1294; Table 1). Marinade retention (%) was significantly higher in birds transported for either 80 km or 160 km compared to those transported for up to 240 km, but non-significant differences were observed among different crating densities and interactions (Table 1). In the present study, sodium chloride was used to marinate the meat, which was reported to increase gelation, water holding capacity, and flavor of meat along with reduction in cooking loss and microbial spoilage (Desmond, 2006). The salt dissociates in water and Cl ions attach to the meat proteins, expanding the lattice of thick and thin filaments due to an increase in the repulsion between negatively charged molecules, and resulting in greater hydration (Warner, 2017). In this experiment, all the birds were exposed to transportation stress; therefore, the distance did not affect marinade uptake (%). Similarly, Woelfel and Sams (2001) compared marination properties of pale (after heat stress) and normal breast meat; they reported non-significant differences for marinade uptake and drip losses, but paler fillets showed a significant increase in cooking losses. Higher marinade retention in 80 and 160 km transportation treatments might be associated with the lower rate of pH fall and relatively higher ultimate pH (Table 1) compared to those subjected to 240 km transportation, which ultimately affects water holding capacity of meat (Caldara et al., 2012). These findings further confirm the findings of Bianchi et al. (2007), who proposed that, with heat stress, the water binding capacity of proteins decreases resulting in lower marinade retention. Nevertheless, the scarcity of data regarding the effect of transportation distances and density on marination properties remains.
Broilers transported for up to 240 km had higher (p<0.0001) shear force (N/cm2) of raw meat (measured after cooking) compared to the other treatments (80 and 160 km), whereas in marinated breast meat, shear force increased gradually (p<0.0001) with each level increase in transportation distances (Table 1). Similarly, the birds transported at crating densities of 12 and 15 birds per crated also had significantly higher (p<0.0001) shear force (N/cm2) for raw breast meat compared to those crated at 10 birds per crate (Table 1). However, non-significant differences were observed in transportation distance × crating density interactions in both types of meat (Table 2). These findings suggest an increase in meat shear force or toughness in birds exposed to higher stress. The birds in the 240 km transportation treatment had the longest exposure to heat stress, which increased drip loss, decreased muscle water contents, and increased toughness of the meat (Lu et al., 2010). Studies have reported a positive relation between muscle water contents and meat tenderness (Silva et al., 2011). Therefore, the decrease in intermuscular water content may result in increase in the shear force required to cut meat. A similar increase in meat shear force in broilers after transportation has been previously reported (Doktor and Połtowicz, 2009; Yalçin and Guler, 2012). A significant increase in drip loss %, cooking loss %, and shear force was also reported in Boer goats transported at higher crating density compared to those transported at lower crating density and without transportation treatment (Nikbin et al., 2016).
Meat lightness, redness, and chroma observed at 2 h PM were not affected by transportation distances, crating densities, and their interactions (p>0.05), while meat yellowness and hue (observed at 2 h PM) were significantly higher in birds transported for up to 80 km and lower in those transported for up to 240 km (Table 3). The increase in meat yellowness could be due to the induction of stress in birds subjected to transportation compared to those slaughtered on a farm. A rapid PM pH decline as described above has been reported to induce pale, soft, and exudative (PSE) or similar conditions in meat, characterized with increased yellowness (b*) of meat imparting a pale color (Warriss, 2010; Wilhelm and Maganhini, 2010). At 24 h PM, meat lightness (L*) increased significantly with the increase in transportation distances and crating density. Birds transported for up to 240 km and those crated at 15 birds per crate density had significantly higher meat L* compared to the other treatments (Table 3). The heat stress due to higher crating density increased the light scattering (high L* value) due to loosening of muscle structure under low pH (Wang et al., 2017). Moreover, no effect (p>0.05) of different transportation distances, crating density, and their interactions on 24 h PM meat redness, yellowness, and hue were observed. Zhang et al. (2019) also reported an increase in lightness (L*) and decrease in a* and b* of the broiler breasts reared in summer compared to those reared in winter. Bianchi et al. (2006) compared different transportation distances (i.e., <40, 40–210, and >210 km) in Italy and reported that the increase in transportation distance resulted in a significant decrease in meat redness and had no effect on lightness and yellowness.
| Parameters | Distance | Cratingdensity | P value | ||||||
|---|---|---|---|---|---|---|---|---|---|
| 80 km | 160 km | 240 km | Low | Medium | High | Distance | Density | Interactions | |
| L* | 55.79±0.59b | 54.79±0.54b | 58.28±0.59a | 55.29±0.64b | 56.24±0.69ab | 57.33±0.64a | 0.0002 | 0.0391 | 0.4295 |
| a* | 14.17±0.47 | 15.19±0.31 | 14.76±0.38 | 14.39±0.46 | 14.64±0.33 | 15.10±0.40 | 0.2006 | 0.4436 | 0.5082 |
| b* | 17.96±0.65 | 18.54±0.44 | 16.76±0.41 | 17.47±0.45 | 17.79±0.66 | 18.00±0.50 | 0.0670 | 0.7786 | 0.6518 |
| c | 22.99±0.53ab | 24.09±0.36a | 22.37±0.45b | 22.69±0.47 | 23.20±0.53 | 23.56±0.45 | 0.0411 | 0.4284 | 0.6972 |
| h | 51.56±1.54 | 50.59±1.02 | 48.62±0.84 | 50.55±1.11 | 50.30±1.40 | 49.92±1.10 | 0.2234 | 0.9320 | 0.4087 |
Cratingdensity: Low=10 birds/crate (0.050 m2/bird), 12 birds/crate (0.042 m2/bird), and 15 birds/crate (0.033 m2/bird)
L*=lightness, a*=redness, b*=yellowness, c=chroma, h=hue
This study showed that transportation of broilers is a stressful process and that the negative impacts of transportation on meat quality increase with the increase in transportation distance and crating density. Transportation distances above 80 km and crating densities above 12 birds per crate resulted in significant increases in meat losses. However, keeping birds at lower crating densities during long journeys can reduce the negative impacts on broilers. The worldwide increase in summer temperatures accentuates the need to devise strategies to reduce the exposure of birds to high temperatures during transportation.
Authors certify that there is no conflict of interest.