原著論文
ADP-グルコーストランスポーター遺伝子の新規アリルの同定と複数の機能性成分を高含有するオオムギ品種育成への利用
2018 年 20 巻 2 号 p. 124-132

詳細
2018 年 20 巻 2 号 p. 124-132
オオムギは穀粒にβ-グルカンなどの機能性成分を多く含む特徴をもつが,その含量をさらに高めたオオムギ品種が求められている.本研究ではADP-グルコーストランスポーターをコードするLys5遺伝子の変異がβ-グルカンなど複数の機能性成分の含量を著しく向上させることを報告する.「四国裸97号」の変異集団から選抜された「谷系QM-1」と,「谷系QM-1」に農研機構中央農業研究センター北陸研究拠点で育成された系統を交配して得られた後代系統の一部では,穀粒の粒厚が薄く,表面にしわがあった.これらのしわ粒系統では総デンプン含量が著しく低下しており,大粒デンプンの形態異常が観察された.また,β-グルカンに加えてアラビノキシラン,フルクタン,レジスタントスターチ,γ-アミノ酪酸を著しく多く含んでいた.Lys5遺伝子の変異アリルをもちβ-グルカン含量が高い既存品種「ビューファイバー」においてもこれらの機能性成分含量の増加や,デンプン粒の形態異常が観察された.そこで「谷系QM-1」とそれを母本とする後代系統についてLys5遺伝子の塩基配列を解析したところ,しわ粒系統のみに共通する1カ所の非同義置換が見出されたことから,この変異が「谷系QM-1」において複数の機能性成分が増加した原因と考えられた.「谷系QM-1」の新規lys5アリルは様々な機能性成分を多く含むオオムギ品種の育成に有用であると考えられたため,アリル名をlys5.iとし,遺伝子型判別用のdCAPSマーカーを開発した.
日本では古くからオオムギを食用に利用してきた(Baik and Ullrich 2008).現在はオオムギ食の血中コレステロールや血糖値の低下,満腹感の持続によるエネルギー摂取量の低減,内臓脂肪蓄積の抑制などの効果が実証され,戦後の急速な食生活の変化により年々患者が増え続けている生活習慣病の予防と改善におけるオオムギ食の価値が見直されている(青江 2015).これまでオオムギの機能性成分として,(1-3,1-4)-β-Dグルカン(以下,β-グルカン)が注目されてきたが(青江 2015),近年はフルクタンやレジスタントスターチなどβ-グルカン以外の機能性成分にも注目が集まりつつある(妹脊 2016).このような状況の中,健康機能性を向上させたオオムギ品種の開発が求められている.
β-グルカンに代表される水溶性食物繊維には腸内細菌による発酵を促進し,腸内環境を調整する効果がある(Lattimer and Haub 2010).それに加え,β-グルカンには冠状動脈心疾患の予防効果があり,アメリカやカナダ,欧州連合でオオムギ食品のヘルスクレーム(健康強調表示)が認証されており(青江 2011,2015,European Food Safety Authority 2009,U.S. Food and Drug Administration 2006),日本でも大麦β-グルカンを関与成分とした製品が機能性表示食品として12件届出されている(2018年7月20日現在,http://www.caa.go.jp/policies/policy/food_ labeling/about_foods_with_function_claims/).また,日本人を対象としたβ-グルカンを含むオオムギと含まないオオムギを用いた摂取試験においても,腹部の内臓脂肪面積が100平方cm以上の群ではβ-グルカンを含むオオムギの摂取による有意な体重,胴囲,BMI,内臓脂肪の減少効果が実証されている(Aoe et al. 2017).
オオムギ穀粒に含まれるβ-グルカン以外の食物繊維や機能性多糖として,胚乳細胞壁のβ-グルカンに次ぐ主要構成物であるアラビノ-(1,4)-β-D-キシラン(以下,アラビノキシラン),スクロースにフルクトースが重合した構造のフルクタン,消化酵素による消化を受けにくいデンプンであるレジスタントスターチなどがあり,これらの成分にもコレステロールや血糖値の低減作用があることが報告されている(Bernstein et al. 2013, Knudsen 2014, Lattimer and Haub 2010, Ritsema and Smeekens 2003).フルクタンにはミネラル吸収の促進作用も報告されている(Lattimer and Haub 2010).γ-アミノ酪酸(GABA)は哺乳動物の中枢神経系において主に抑制系の神経伝達物質としてはたらくアミノ酸であり,血圧上昇抑制効果が知られている(Takahashi et al. 1961).オオムギ穀粒にもGABAが含まれており,特に高リジン変異体は高濃度のGABAを含んでいることが報告されている(Nogata et al. 2012).
これまでにβ-グルカンの機能性に着目した「ビューファイバー」(塔野岡ら 2011),「ワキシーファイバー」(栁澤ら 2016)が育成されている.これらの品種ではデンプン合成の基質であるADP-グルコースをアミロプラストに輸送するADP-グルコーストランスポーターをコードするLys5遺伝子に変異があり,デンプン含量が減少し,β-グルカン含量が著しくする増加する特徴をもつ(Munck et al. 2004,Patron et al. 2004,塔野岡ら 2011).
海外では機能性成分をセールスポイントとするオオムギ品種「Himalaya 292(後のBARLEYmaxTM)」がオーストラリアで育成され,スーパー大麦として日本でも輸入・販売されている(妹脊 2016).「Himalaya 292」は原品種「Himalaya」の高アミロース変異体として単離され,デンプン合成酵素starch synthase IIaのナンセンス変異が原因である(Morell et al. 2003).「Himalaya 292」にはデンプン含量が低く,β-グルカン含量が高いことに加えて,レジスタントスターチ含量も高い特徴がある(Bird et al. 2004,妹脊 2016).
近年の健康ブームにより,国内ではβ-グルカンをより多く含み麦ご飯の食味が優れるモチ性オオムギの開発が進められてきた(前島 2016,関ら 2018,Yanagisawa et al. 2011).今後,さらなるオオムギの需要拡大のためには,「Himalaya 292」のようにβ-グルカン含量に加えて他の機能性成分含量も高めることにより高付加価値化を実現したオオムギ品種の開発が重要である.本研究では,「谷系QM-1」とそれを母本とする後代系統がβ-グルカンに加えてフルクタンやGABAなど複数の機能性成分を非常に多く含むことを明らかにした.またこれらの機能性成分を高含有する国産オオムギ品種の育成促進のために,「谷系QM-1」の機能性成分増加の原因遺伝子変異を明らかにし,遺伝子型判別のためのDNAマーカー開発を行った.
「ビューファイバー」,「四国裸97号」,「谷系QM-1」の機能性成分の分析には,農研機構次世代作物開発研究センター(茨城県つくば市)において栽培・収穫した穀粒を用いた.なお,「谷系QM-1」はモチ性の二条裸麦「四国裸97号」のアジ化ナトリウム処理により作出した系統である(一ノ瀬ら 2015).「谷系QM-1」および「ビューファイバー」を母本とする後代系統,「ファイバースノウ」,「はねうまもち」については,農研機構中央農業研究センター北陸研究拠点(新潟県上越市)において栽培・収穫した穀粒を用いた.栽培条件の詳細を表1にまとめた.
| 産地 | 試験名 | 試験年度 | 播種日 | 播種法 | 畦幅 (cm) |
株間 (cm) |
基肥(成分kg/10 a) | 追肥(成分kg/10 a) | ||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 施用日 | N | P | K | 施用日 | N | K | ||||||||
| 新潟県上越市 | 点播 | 2016 | 9月27日 | 二条千鳥点播 | 75 | 13 | 9月27日 | 6.2 | 2.2 | 5.3 | 2月27日 | 7.0 | 7.0 | |
| ドリル播 | 2016 | 10月13日 | 4条ドリル播(1区2条) | 75 | ― | 10月7日 | 6.2 | 2.2 | 5.3 | 3月1日 | 7.0 | 7.0 | ||
| 茨城県つくば市 | 点播1) | 2016 | 11月9日 | 点播 | 70 | 8.5 | 11月9日 | 5.0 | 7.5 | 5.0 | なし | |||
1) 開花後3週目より,雨避け栽培.
穀粒をカミソリで横断し,走査型電子顕微鏡(S-3400N,日立ハイテクノロジーズ)を用いて種子横断面のデンプン粒を無処理,低真空条件(80 Pa, 8 kV)で観察した.
3. 機能性成分の定量原麦を粉砕機(Cyclotec 1093 sample mill, FOSS)により0.5 mm以下に粉砕した全粒粉を用い,乾物重当たりの重量で算出した(GABAを除く).
β-グルカン,フルクタン,レジスタントスターチ含量は,それぞれ「β-Glucan Assay Kit (Mixed Linkage)(AOAC Method 995.16)」,「Fructan Assay Kit(AOAC Method999.03)」,「Resistant Starch Assay Kit(AOAC Method 2002.02)」(全てMegazyme)を用いて測定した.
総デンプン含量は80% エタノールで2回洗浄して可溶性糖を除いた全粒粉について「Total Starch Assay Kit (AA/AMG)」(Megazyme)を用い,レジスタントスターチを含むサンプルに適したKOH法とAOAC Method 996.11を組み合わせて測定した.
アラビノキシラン含量は以下の方法で測定した.全粒粉20 mgを99.5% エタノール中で100℃,5分間加熱して内在性の酵素を失活させた.試料に含まれるアラビノガラクタンを除去するために蒸留水を加えて50% エタノールとし,15分間振盪後,遠心分離(1000 × g,10分)し,上清を除いた.沈殿に50% エタノールを加えて懸濁後,再度遠心分離(1000 × g,10分)し,上清を除いた.沈殿に0.5 mLの12 M硫酸を加えて室温で30分間振盪し,5.5 mLの蒸留水を加えて100℃で2時間加熱した後,遠心分離(1000 × g,10分)した.この上清をA液とする.A液150 μLに100 μLの蒸留水,500 μLの0.2 M Tris-HCl(pH 8.6),45 μLの20 mg/mL NAD,7.5 μL(105 mU)のβ-ガラクトースデヒドロゲナーゼS(ロシュ)を加えて40℃,1時間反応し,酵素を加えないものをブランクとした340 nmの吸光度からアラビノース含量を求めた.また,A液について「D-Xylose Assay Kit」(Megazyme)を用いてキシロース含量を測定した.アラビノース含量とキシロース含量の合計に0.88を乗じてアラビノキシラン含量を算出した.
GABAの抽出,分析は以下の方法で実施した.全粒粉に0.1 M HClを添加し,懸濁,攪拌し,4℃で一晩保温した.懸濁液を遠心分離し(10,000 × g,20分),上清を回収した.さらに沈殿に0.1 M HClを添加後,懸濁,攪拌し,上清を回収する操作を2回繰り返し,合わせた上清を100 mg/mLに定容し,抽出液とした.抽出液10 μLを用いて,桑野ら(1987)に従い,フェニルイソチオシアネートによりラベル化したフェニルチオカルバミル-GABAをHPLCにより分離し,同様にラベル化した標品のピーク面積値を基に定量した.
4.Lys5遺伝子の塩基配列解析葉サンプルから酢酸カリウム法(Dellaporta et al. 1983)の改変法によりゲノムDNAを抽出した.3 cm程度の葉サンプルに550 μLのDNA抽出緩衝液(100 mM Tris-HCl pH 8.0,50 mM EDTA,1.25% SDS,500 mM NaCl,7% エタノール)を加えて破砕し,65℃で10分保温した後,200 μLの5 M酢酸カリウムを加え,遠心分離(15,000 × g,10分)した.上清を回収し,等量のイソプロパノールを加え,遠心分離(15,000 × g,10分,4℃)した.上清を除き,沈殿を70% エタノールでリンスした後,300 μLの1.5 mg/ml RNaseを含むTE溶液に溶解した.Lys5遺伝子を増幅するためのPCR反応液組成は滅菌蒸留水12.2 μL,5 × PCR緩衝液4 μL,dNTP mix(各2.5 mM)1.6 μL,フォワードプライマー(5′-AGGACT CAAGAACTCCTAGG-3′)(10 μM)0.4 μL,リバースプライマー(5′-CAATTGCGCTAGGCTCTACA-3′)(10 μM)0.4 μL,ゲノムDNA 1.0 μL,PCR酵素(PrimeStar GXL, Takara)0.4 μLとした.PCR反応は最初に98℃ 30秒,続いて98℃ 10秒,65℃ 15秒,68℃ 30秒を35サイクル,最後に68℃ 1分とした.増幅されたDNA断片をQuantum Prep PCR Kleen Spin Columns(Bio-Rad)を用いて精製し,塩基配列を依頼分析(ファスマック)により解析した.
「谷系QM-1」の種子表面にはしわがあり(以下,しわ粒とする)(図1A),「四国裸97号」と比較して著しく粒厚が薄かった.完熟粒の穀粒横断面のデンプン粒の形態を走査型電子顕微鏡で観察したところ,「四国裸97号」の大粒デンプンは円盤状であり,1本の赤道周りの溝が見られたのに対し,「谷系QM-1」の大粒デンプンには厚みがあり表面の著しい起伏が見られた.また,赤道周りの溝が複数本あり,かつ溝が湾曲している大粒デンプンも観察された(図1B).

供試材料の原麦外観とデンプン粒の形態.
(A)原麦外観.(B)デンプン粒の電子顕微鏡観察像.大粒デンプンの赤道まわりの溝を白矢印で示した.赤道まわりの湾曲した溝を黒矢印で示した.スケールバーは10 μm.
「谷系QM-1」のβ-グルカン含量は約17%であり,「四国裸97号」の約2.5倍,「ビューファイバー」の約1.3倍であった(図2).反対にデンプン含量は「四国裸97号」(65%),「ビューファイバー」(46%),「谷系QM-1」(35%)の順であった.アラビノキシラン,フルクタン含量も「谷系QM-1」,「ビューファイバー」で増加しており,「谷系QM-1」の方がより多く含んでいた.レジスタントスターチは「谷系QM-1」,「ビューファイバー」で同程度に増加していた.

機能性成分の含量.
乾物重当たりの各成分の含量を%で示した.GABAはmg/100 gで示した.つくば市のハウス,上越市の点播およびドリル播栽培のサンプルをそれぞれグレー,青,オレンジのバーで示した.「通常」は通常粒系統,「しわ」はしわ粒系統を示す.値は3~4反復の平均値,エラーバーは標準偏差.穀粒サンプルが十分に得られなかった試験区は欠測.
「谷系QM-1」と「ビューファイバー」におけるβ-グルカン,アラビノキシラン,デンプン含量の増減は一ノ瀬ら(2015)と同様の傾向であった.
2. 「谷系QM-1」を母本とする後代系統の育成と特性β-グルカン含量が高い寒冷地適応品種を育成するために,「谷系QM-1」と寒冷地適応系統である「北陸皮48号」あるいは「北陸皮49号」を交配し,栽培性や収量性の改良を指標として二条裸麦の「新系GNT237W」(モチ),「新系GNT238」(ウルチ),「新系GNT239」(ウルチ),「新系GNT275W」(モチ),二条皮麦の「新系GT273W」(モチ)を育成した.また,「ビューファイバー」と「北陸皮49号」の交配により二条裸麦「新系GNT232」(ウルチ)を育成し,比較として用いた.モチ性系統には系統名に「W」を付した.これらの系統のうち「新系GNT238」以外の系統は「谷系QM-1」や「ビューファイバー」と同様にしわ粒であった(図1A).
デンプン粒の観察から,しわ粒系統では「谷系QM-1」と同様に大粒デンプンの形態異常が見られたが,通常粒の「新系GNT238」には見られなかった(図1B).
点播(系統養成)とドリル播(生産力検定)の2条件で栽培した穀粒サンプルを用いて,総デンプン,β-グルカン,アラビノキシラン,フルクタン,レジスタントスターチ,GABA含量を測定した.「谷系QM-1」を母本とする後代のしわ粒系統のデンプン含量は「新系GNT238」や標準品種である六条皮麦の「ファイバースノウ」等と比較して約半分であった(図2).β-グルカン含量は「新系GNT238」,「ファイバースノウ」の約3倍,β-グルカン含量がやや高いモチ性品種である「はねうまもち」と比較しても2倍以上であった.また,フルクタン含量は「ファイバースノウ」と比較して約4倍,レジスタントスターチ含量は約1.5倍であった.アラビノキシラン含量は「ファイバースノウ」の約1.2倍であったが,皮麦は裸麦よりも全粒粉中のアラビノキシラン含量が高いことが報告されており(Holtekjølen et al. 2006),本研究でも皮麦である「新系GT273W」のアラビノキシラン含量が最も高かった(図2).そこで裸麦同士で比較するとしわ粒の「新系GNT237W」,「新系GNT239」,「新系GNT275W」は「新系GNT238」の2倍以上のアラビノキシランを含有していた.さらにしわ粒系統は「ファイバースノウ」の15~30倍のGABAを含有していた.
「谷系QM-1」を母本とする後代のしわ粒系統のβ-グルカン,フルクタン,アラビノキシラン含量は「ビューファイバー」を母本とする後代のしわ粒系統「新系GNT232」よりも高かったが,デンプン,レジスタントスターチ含量は「新系GNT232」の方が高かった.
いずれの成分についても点播およびドリル播での含量の差異は小さかった.
3. 「谷系QM-1」の機能性成分増加に関する原因遺伝子の解析「谷系QM-1」とそれを母本とする後代のしわ粒系統に共通して観察されたデンプン含量の減少,穀粒表面のしわ,大粒デンプンの形態異常は,lys5.hアリルをもつ「ビューファイバー」や他のlys5変異体の観察結果と類似していた(図1,図2)(Patron et al. 2004).そこで,「谷系QM-1」とそれを母本とする後代系統のLys5遺伝子の開始コドンから終始コドンまでのイントロンを含めた約1500 bp領域の塩基配列を解析したところ,「谷系QM-1」とそれを母本とする後代のしわ粒系統に共通する1カ所の塩基置換を第3エクソン内に見出した(図3).この塩基置換によりLys5遺伝子にコードされるADP-グルコーストランスポータータンパク質の269番目のセリン(S269)がアスパラギンに置換されていた(S269N変異).S269N変異は原品種である「四国裸97号」および通常粒系統「新系GNT238」には存在しなかった.「谷系QM-1」のLys5遺伝子の変異は,「ビューファイバー」のlys5.hアリルが有する228番目のプロリンからセリンへの変異(P228S変異)やlys5.e,β-グルカン含量が高くなると報告されているlys5.f,lys5.gアリル(Munck et al. 2004, Patron et al. 2004)とは異なっていたことから,新規変異アリルとしてlys5.iと名付けた.lys5.i変異アリルのGenbankアクセッション番号はLC377180である.

「谷系QM-1」の変異とADP-グルコーストランスポータータンパク質の構造.
(A)Lys5遺伝子の一部のゲノム配列.全ての系統に共通する塩基をアスタリスクで示し,対応するアミノ酸配列をその下に示した.エクソンは大文字,イントロンは小文字で示した.「ビューファイバー」が有するlys5.h,「谷系QM-1」が有するlys5.iの変異塩基およびアミノ酸を白黒反転した.
(B)ADP-グルコーストランスポータータンパク質の構造の模式図.6個の膜貫通領域をN末端側からH1~H6で示した.lys5.hとlys5.iの変異アミノ酸の位置を示した.
lys5.i変異アリルを検出するdCAPSマーカーのPCRおよび制限酵素処理条件を図4AおよびBに示した.フォワードプライマー内のSphIサイトに1塩基のミスマッチを入れてPCR産物がそのSphIで切断されないようにした.PCR反応後,PCR産物をSphIで消化し,3%アガロースゲルで電気泳動した.
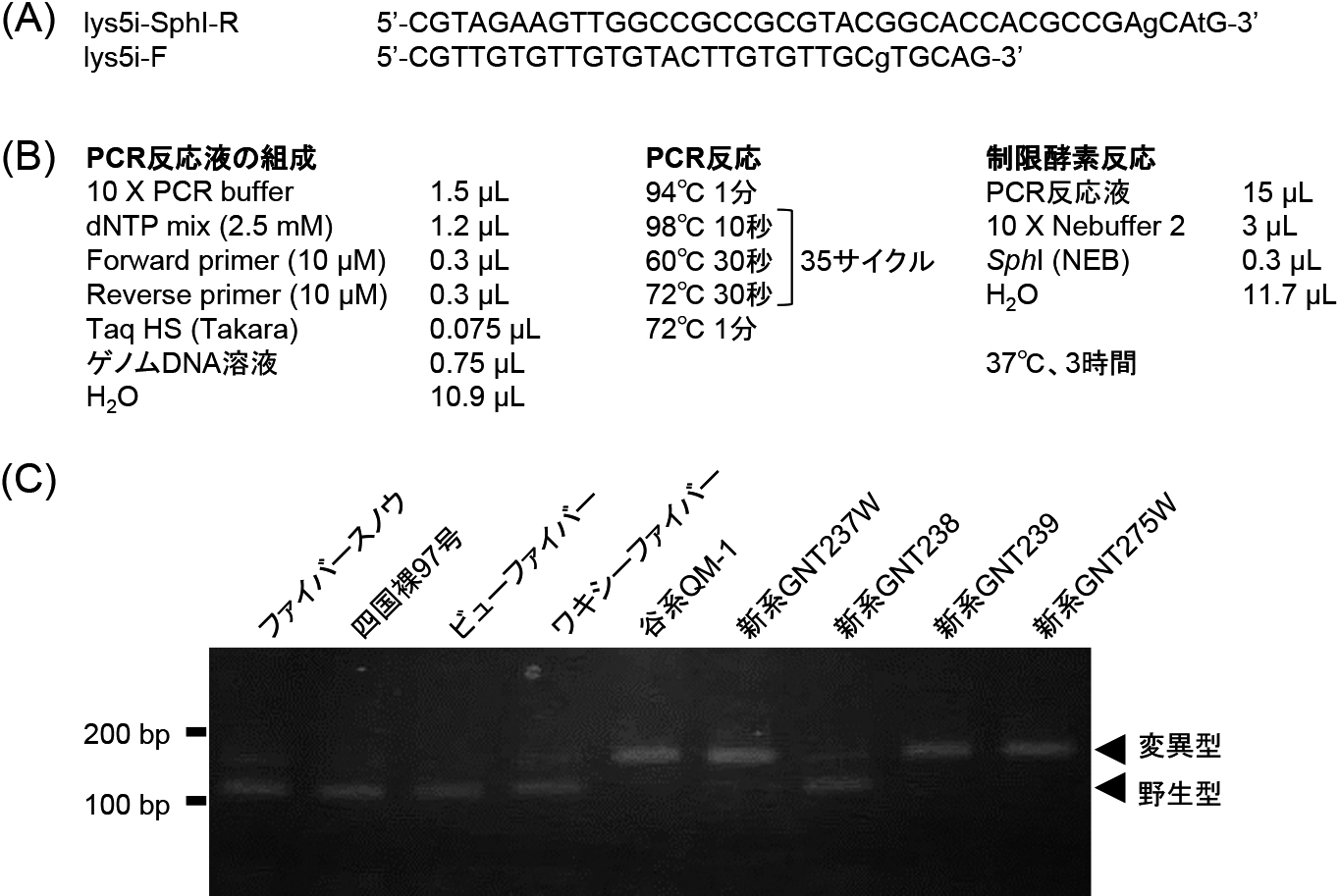
lys5.i変異アリルを検出するdCAPSマーカー.
(A)dCAPSマーカーのプライマー配列.ミスマッチ塩基を小文字で示した.
(B)PCR反応条件と制限酵素処理条件.
(C)電気泳動図.3%アガロースを用い,TAE緩衝液中で電気泳動した.
「谷系QM-1」とそれを母本とする後代のしわ粒系統ではSphIで切断された約120 bpのバンドが見られ,その他の品種・系統では切断されてない160 bpのバンドが見られた(図4C).lys5.i変異アリルをもたない系統でも消化されなかった約160 bpのバンドがわずかに見られるが,遺伝子型の判別には問題なかった.したがって,本マーカーによりlys5.i変異アリルを検出できることが明らかになった.
本研究によりLys5遺伝子の変異アリルをもつ「谷系QM-1」,「ビューファイバー」,およびそれらを母本とする後代系統はβ-グルカンだけでなく,他の複数の機能性成分(フルクタン,レジスタントスターチ,アラビノキシラン,GABA)も多く含むオオムギであることが明らかになった(図2).特にlys5.i変異アリルは効果が高く,最も機能性成分の含量が高い「新系GNT237W」,「新系GNT275W」の機能性成分の合計は,同条件で栽培したlys5.hアリルをもつ「新系GNT232」のそれよりも高かった.これらの系統ではβ-グルカンが「ファイバースノウ」の3倍以上と著しく高い一方で,レジスタントスターチの割合は相対的に低かった.これはβ-グルカン含量と比較して,レジスタントスターチの割合が高い「BARLEYmaxTM」の特徴とは異なっており(妹脊 2016,Topping et al. 2003),原因遺伝子の違いを反映している.
機能性成分の中でβ-グルカンは胚乳に多く含まれるのに対し,アラビノキシランは果皮や糊粉層などのふすま部分に,フルクタンは胚部分に多いため(Bernstein et al. 2013),搗精による損失が大きい.「谷系QM-1」とそれを母本とする後代系統の「新系GNT237W」,「新系GNT239」,「新系GNT275W」は裸麦であり,搗精せずに全粒粉として利用できるため,アラビノキシランやフルクタンを損失なく摂取できる点が機能性食品素材として優れている.
2. 機能性成分の増加「谷系QM-1」やそれを母本とする後代のしわ粒系統では胚乳容積が減少したことにより,果皮やアリューロン層など胚乳以外の組織の乾物重当たりの量が相対的に多くなる.今回分析した機能性成分の中で胚乳以外の組織に多く含まれるアラビノキシランやフルクタンについては,合成量の増加と胚乳容積減少により相対的含量が増大したことの両方が含量の増加に寄与していると考えられる.
「谷系QM-1」を母本とする後代のしわ粒系統は8~10%のフルクタンを含んでいた(図2).これは外国産オオムギやコムギの0.5~4%,および,親系統である「谷系QM-1」(約5%)と比較して著しく高い値である(Huynh et al. 2012, Nemeth et al. 2014).一般にコムギでは寒冷地適応品種では茎葉にフルクタンを高濃度に蓄積しやすいことが報告されており(湯川・渡辺 1995),オオムギでも同様の傾向のデータが得られている(中田ら 未発表データ)ことから,寒冷地適応系統である「北陸皮48号」や「北陸皮49号」との交配で得られた「谷系QM-1」の後代しわ粒系統はLys5遺伝子の変異の効果に加え,フルクタンを蓄積しやすい遺伝的背景である可能性が考えられる.また,積雪量の多い上越市で栽培したことが影響した可能性も考えられる.
Nogata et al.(2012)は高リジン変異体に由来するlys3.aやlys5.hアリルをもつオオムギ系統では,リジンだけではなくGABAの前駆体であるグルタミン酸を含む多くの遊離アミノ酸が増加しており,GABAも増加していることを報告している.今回解析したしわ粒系統でもグルタミン酸など遊離アミノ酸の増加が検出された.しかし,GABAや遊離アミノ酸の増加とLys5遺伝子変異が影響すると思われる糖代謝の関係について現時点では不明である.
モチ性オオムギはウルチ性オオムギと比較してβ-グルカン含量が高い(前島 2016,関ら 2018,Ullrich et al. 1986,Yanagisawa et al. 2011).本研究で育成したモチ性の「新系GNT237W」,「新系GT273W」,「新系GNT275W」ではβ-グルカンとアラビノキシラン含量がウルチ性の「新系GNT239」よりも高かった(図2).一方でフルクタンとGABA含量はウルチ性の「新系GNT239」の方が高い.このことから機能性成分によってモチ性による影響が異なることが示唆されるが,分析に用いた系統数が少ないことから今後さらなる検討が必要である.また,共にウルチ性であるがlys5.iをもつ「新系GNT239」とlys5.hをもつ「新系GNT232」を比較した場合,β-グルカンとアラビノキシラン含量は同程度であるが,フルクタンとGABA含量は「新系GNT239」の方が高いことから,前述の通りlys5.iアリルの方が機能性成分含量に対する効果が大きいと考えられる.
β-グルカン,アラビノキシラン,レジスタントスターチ含量はつくば市と上越市(点播およびドリル播)の3条件で,フルクタンとGABAは上越市の2条件で栽培した穀粒で同様に増加していた.このことからLys5遺伝子変異による複数の機能性成分含量を増加させる効果は栽培条件の変化による増減よりも大きいと考えられる.
3. ADP-グルコーストランスポーターの機能への影響ADP-グルコーストランスポータータンパク質には6個の膜貫通領域(H1~H6)があり,H1,H3,H5は折れ曲がった構造を取っている(図3B).「谷系QM-1」のlys5.iアリルでアスパラギンに置換されていたS269はH4に位置しており,トウモロコシのADP-グルコーストランスポーター(Brittle-1)や,構造のよく似たウシのミトコンドリアATPキャリアータンパク質にも保存されていた(Pebay-Peyroula et al. 2003, Sullivan et al. 1991).このことはS269がADP-グルコーストランスポータータンパク質の構造と機能に非常に重要なアミノ酸であることを示唆しており,S269N変異により機能に欠陥を生じると考えられる.一方,「ビューファイバー」のlys5.hアリルでセリンに置換された228番目のプロリンはH3の折れ曲がり構造に重要なアミノ酸であることから(Patron et al. 2004),ADP-グルコーストランスポータータンパク質の構造と機能に欠陥を生じると推測できる.デンプンや機能性成分含量への影響の程度から,「ビューファイバー」のP228S変異より「谷系QM-1」のS269N変異の方がタンパク質機能に重大な影響を及ぼしていると考えられる.
ADP-グルコーストランスポーターの機能低下によりデンプン合成の低下と,胚乳細胞質でのADP-グルコースの過剰蓄積が起こる(Patron et al. 2004).Munck et al.(2004)はデンプン合成が低下するとβ-グルカンなど他の炭水化物の合成を増加することで総炭水化物含量が維持されるとしている.本研究のしわ粒系統においても,デンプン合成に使用されなかった過剰な糖がβ-グルカンやフルクタン等の多糖類合成の基質として消費された結果,機能性多糖を高濃度に含む穀粒になったと考えられる.
4. 今後の展望と課題「谷系QM-1」の複数の機能性成分を含有する特徴が同系統を母本とする後代のしわ粒系統で維持されていたことから,「谷系QM-1」は機能性成分を高含有する国産オオムギ品種育成の交配母本として非常に有用である.本研究で原因遺伝子変異を特定し,DNAマーカーを開発したことにより,機能性成分を高含有する系統の選抜が可能となり,今後の品種育成が加速される.一方,「谷系QM-1」とそれを母本とする後代系統はデンプン含量が非常に低いことからしわ粒となり,千粒重も30 g程度と二条大麦としては非常に低い(データ略).粒重に加えて整粒歩合,収量性,耐雪性,出芽性などの品質や栽培性については改善の余地があり,育種的に解決すべき今後の課題である.また,β-グルカンやフルクタン含量が産地の違いにより変動したことから,これらの機能性成分含量の高位安定化技術にも取り組む必要がある.
本研究を遂行するにあたり,農研機構中央農業研究センター北陸業務科技術専門職員には圃場管理業務や収穫物調整業務でご尽力いただいた.作物開発研究領域畑作物育種グループ契約職員には品質分析や育種業務全般補助に多大なる貢献をいただき感謝の意を表する.
本研究の一部はJSPS科研費JP18K05604の助成を受けて実施した.材料の一部は農業生物資源ジーンバンクより提供を受けた.