3. Results
3.1. Phylogeny
The ITS-LSU phylogenies were completed when the best likelihood values -InL = 28647.12 for the optimized tree in RAxML and -InL = 28422.11 and 28431.56 for cold chains in MrBayes. The ML and BI phylograms did not show the topology conflict having the supported branches (i.e., MLBS < 50, BIP < 0.90) (Fig. 1). The phylogenetic tree showed closely relationships between /resudial clade, /phlebioid clade, and Candelabrochaete africana Boidin (MLBS/BIP = 82/0.99) within the order Polyporales (MLBS/BIP = 100/1), corresponding to the previous studies (Binder et al., 2013; Justo et al., 2017). Although all Hypochnicium s. lat. species grouped in /residual clade, many species were nested within the genera Abortiporus, Bulbillomyces, Cymatoderma, Podoscypha, and Pouzaroporia. Hypochnicium bombycinum, the type species of Hypochnicium, formed monophyletic clade (MLBS/BIP = 89/0.98) with H. multiforme (Berk. & Broome) Hjortstam, H. lyndoniae (D.A. Reid) Hjortstam, and Hypochnicium sp. WY-DT1. The genus Gyrophanopsis (Gy. polonensis and Gy. japonica) formed monophyletic clade with strong support (MLBS/BIP = 100/0.99) as sister clade of Bulbillomyces farinosus (Bres.) Jülich (MLBS/BIP = 94/1). The remaining Hypochnicium species formed monophyletic clade with moderate support (MLBS/BIP = 72/0.99); namely, H. michelii Tellería, M. Dueñas, Melo & M.P. Martín, H. geogenium (Bres.) J. Erikss., H. pini, H. punctulatum, H. sphaerosporum, H. subrigescens, H. wakefieldiae, and two new species. This monophyletic clade of Hypochnicium s. str. positioned apart from Gyrophanopsis clade.

Fig. 1 - Maximum likelihood tree based on the LSU + ITS sequences of Hypochnicium (s. lat.) species. Each value on branch shows maximum likelihood bootstrap value (≥ 60) and Bayesian inference posterior probability (≥ 0.90). Hypochnicium s. lat. and Gyrophanopsis were shown with blue and orange characters, respectively.
In the ITS dataset, the best likelihood values were -InL = 9179.27 in RAxML and -InL = 9406.76 and 9408.01 in MrBayes, respectively. Both analyses demonstrated the similar topology (Fig. 2). The ITS phylogenies supported each monophyletic lineage of Hypochnicium s. str. (MLBS/BIP = 88/0.99), Gyrophanopsis (MLBS = 99/1), and the remaining Hypochnicium species (MLBS/BIP = 68/0.90). At least 14 species of Hypochnicium were included in “Neohypochnicium” clade: namely, H. albostramineum, H. aotearoae B.C. Paulus, H. Nilsson & Hallenb., H. cremicolor (Bres.) H. Nilsson & Hallenb., H. cystidiatum Boidin & Gilles, H. geogenium, H. guineense Tellería, M. Dueñas, Melo & M.P. Martín, H. huinayense Tellería, M. Dueñas & M.P. Martín, H. michelii, H. microsporum G. Gruhn, Schimann & M. Roy, H. patagonicum Gorjón & Hallenb., H. pini, H. punctulatum, H. subrigescens, and H. wakefieldiae. All Japanese lineages excepting one species, “Neohypochnicium asiaticum”, formed well-supported monophyletic clade (MLBS ≥ 95, BIP ≥ 0.95) with foreign strains of conspecific taxa. On the complete sequences of ITS region (ITS1-5.8S-ITS2), the interspecific variations among “N. asiaticum”, “N. albostramineum” and “N. wakefieldiae” were relatively smaller (2.1-6.3%; 12-35 bps of differences within 535-537 bps) than those among other species of Hypochnicium s. lat. (≥ 3.5%) and “N. asiaticum” showed paraphyletic clade; however interspecific variations among the three taxa exceed those of intraspecific variation within each single species (≤ 1.4%). Also, the RAxML phylogeny including ITS sequence of only three groups indicated each monophyly of “N. wakefieldiae” (MLBS = 100), “N. albostramineum” (MLBS = 99) and “N. asiaticum” (MLBS = 71) (Supplemental Fig. S1); thus, we treated these three groups as different species.

Fig. 2 - Bayesian tree based on ITS sequences of Hypochnicium s. lat. Each value on branch shows Bayesian inference posterior probability (BPI) and maximum likelihood bootstrap value (MLBS). BIS ≥ 0.80 and MLBS ≥ 60 were shown on branches and BIS ≥ 0.95 and MLBS ≥ 90 were shown with bold branches. Red sequences indicate Japanese taxa in this study.
Neohypochnicium N. Maek. & R. Sugaw., gen. nov.
MycoBank No.: MB 842573.
Diagnosis: Basidiomata resupinate, effused, adnate; hymenial surface smooth to odontoid. Hyphal system monomitic; hyphae hyaline, with clamp connections at all septa. Cystidia often present, aseptate, thin- to thick-walled, enclosed or projecting beyond the hymenial surface. Basidia subclavate to suburniform, normally 4-sterigmate, with a basal clamp. Basidiospores ellipsoid to subglobose, smooth, finely verrucose or finely echinulate, thick-walled (not less than 0.5 µm thick), cyanophilous, inamyloid.
Type species: Neohypochnicium perlongicystidiosum N. Maek., Kogi & Norikura.
Gene sequence ex-holotype: LC663690 (LSU), LC663677 (ITS).
Etymology: “Neohypochnicium” [neo (= new) + Hypochnicium] refers to having morphologically similar features to Hypochnicium.
Hypochnicium J. Erikss., Symb. Bot. Upsal. 16: 100 (1958) emend. N. Maek.
Basiodiomata resupinate, effused, adnate. Hymenial surface smooth, grandinioid to odontoid. Hyphal system monomitic; hyphae hyaline, with clamp connections at all septa. Cystidia often present, aseptate, thin- to thick-walled, enclosed or projecting beyond the hymenial surface. Basidia clavate to suburniform, normally 4-sterigmate, with a basal clamp. Basidiospores ellipsoid to subglobose, smooth, thick-walled (not less than 0.5 µm thick), cyanophilous, inamyloid.
Remarks: Hypochnicium emended is primarily characterized by producing smooth, thick-walled (not less than 0.5 µm thick) and cyanophilous basidiospores, clavate to suburniform basidia, and hyphae with clamp connections at all septa. This genus shares these morphological features with Bulbillomyces and Gyrophanopsis. But Bulbillomyces distinguishes from Hypochnicium in that produces sclerotia (Aegerita state) associated with teleomorph. Gyrophanopsis differs from Hypochnicium in that has heavily encrusted septocystidia and only slightly thick-walled basidiospores (less than 0.5 µm thick).
3.2.2. New species
Neohypochnicium asiaticum N. Maek. & Kogi, sp. nov. (Figs. 3A, 4, 7A, 8A)
MycoBank No.: MB 842561.
Diagnosis: Neohypochnicium asiaticum is characterized by having smooth hymenophores, cylindrical to subfusiform cystidia (52-170 × 9-13 µm), and broadly ellipsoid, finely verrucose, thick-walled (0.5-1 µm thick) basidiospores measuring 7-8(-8.5) × 5.5-6.5 µm.
Holotype: TUMH 61220, on decayed wood of coniferous tree, Kokoge, Tottori City, Tottori Pref., Japan, 8 Nov 1983, collected by N. Maekawa. Ex-holotype culture: TUFC 31698. GenBank: LSU = LC663689; ITS = LC663669.
Etymology: “asiaticum” refers to Asia, where the vouchered specimens were collected.
Description: Basidiomes resupinate, adnate, effused, 100-300 µm thick; hymenial surface white, “Buff” to “Pale Luteous” when drying, smooth, sometimes minutely cracked under the lens (×10); margin concolorous with the hymenial surface, thinning out, indeterminate. In vertical section, subhymenium subhyaline, composed of densely interwoven hyphae; subiculum subhyaline, composed of loosely interwoven hyphae at first and then gradually becoming dense. Hyphal system monomitic; hyphae 3-7.5 µm wide, smooth, thin- to thick-walled (up to 1.5 µm thick), with clamp connection at all septa. Cystidia arising from subicular or subhymenial hyphae, cylindrical to subfusiform with obtuse apex, 52-170 × 9-13 µm, smooth, thin- to slightly thick-walled (up to 0.5 µm), with a clamp connection at the basal septum, projecting up to 80 µm beyond the hymenial surface. Basidia clavate to suburniform, 23-42 × 7-8 µm, thin-walled, usually with oily drops, with a basal clamp, producing 4 sterigmata; sterigmata 5-7 µm in length. Basidiospores broadly ellipsoid, 7-8(-8.5) × 5.5-6.5 µm [Lm = 7.6 ± 0.4 µm, Wm = 5.9 ± 0.3 µm, Q = 1.2-1.4, Qm = 1.3 ± 0.1; n=180/9] finely verrucose, thick-walled (0.5-1 µm thick), cyanophilous, non-amyloid.
Characteristics in culture: The optimum growth temperature for the polysporous isolates TUFC 31698, TUFC 31733 and TUFC 100660 was 25 °C. Isolates could grow between 5 and 30 °C, but no visible growth was observed at 35 °C. Growth on MA was 20-26 mm in radius at 25 °C after 5 d in the dark. Mycelial mats at that time white, cottony; reverse side of mycelial mats not discolored; agar not bleached; odor not noticeable. Aerial hyphae 2-6 µm wide, subhyaline, thin- to slightly thick-walled (up to 0.5 µm thick), with clamp connections at all septa, moderately branched, occasionally encrusted. Marginal hyphae 2-4 µm wide, subhyaline, thin-walled, clamp connections at all septa, sparsely branched. Submerged hyphae dense, 4-7.5 µm wide, subhyaline, thin- to slightly thick-walled (up to 0.5 µm thick), with clamp connections at all septa, relatively much branched. Conidia, chlamydospores and cystidium-like cells not observed after 6 wk of incubation at 25 °C.
Species codes (Stalpers, 1978): 7, (12), 14, 21, 30, 39, 52, 53, 54, 82.
Other specimens and cultures examined: JAPAN. TUMH 60588 on dead and decorticated trunk of a coniferous tree, Hasehiji, Ina City, Nagano Pref., 19 Sep 2013, collected by N. Maekawa (culture: TUFC 100660); TUMH 64607 [as H. punctulatum (TMI 17549)] on decayed wood of Cryptomeria japonica D. Don, Miasa-mura, Kitaazuki-gun, Nagano Pref., 12 Jul 1987, collected by N. Maekawa; TUMH 64605 [as H. punctulatum (TMI 9479)] on decayed wood of Pinus densiflora Sieb. & Zucc., Miasa-mura, Kitaazuki-gun, Nagano Pref., 2 Oct 1987, collected by N. Maekawa; TUMH 64609 on decayed wood of a coniferous tree, Takane-machi, Takayama City, Gifu Pref., 28 Sep, 2015, collected by N. Maekawa; TUMH 61227 [as H. eichleri (TMI 17550)] on decayed wood of C. japonica, Kokoge, Tottori City, Tottori Pref., 17 May 1984, collected by N. Maekawa (culture TUFC 31733); TUMH 64608 [as H. eichleri (TMI 10704)] on decayed wood of P. densiflora, Daisen-cho, Saihaku-gun, Tottori Pref., 28 Oct 1984, collected by N. Maekawa; TUMH 64606 [as H. punctulatum (TMI 17548)] on dead trunk of P. densiflora, Okamasu, Kokufu-cho, Tottori City, 14 Jul 1986, collected by N. Maekawa; TUMH 64610 on dead branch of P. densiflora, Daisen-cho, Saihaku-gun, Tottori Pref., 11 Oct 2015, collected by H. Kogi.
Remarks: Neohypochnicium asiaticum is characterized by smooth hymenial surface, broadly ellipsoid and finely verrucose basidiospores measuring 7-8(-8.5) × 5.5-6.5 µm, and cylindrical to subfusiform cystidia measuring 52-170 × 9-13 µm. Neohypochnicium asiaticum is placed in the H. punctulatum complex designated by Nilsson and Hallenberg (2003), with N. albostramineum, N. huinayense, N. punctulatum and N. wakefieldiae. Phylogenetically, N. asiaticum is very close to N. albostramineum, but basidiospores (8-9.5 × 6.5-7.5 µm, Tellería, Dueñas, Melo, Hallenberg, & Martín, 2010) of H. albostramineum are larger than those of N. asiaticum. Neohypochnicium asiaticum also resembles N. wakefieldiae having broadly ellipsoid to subglobose basidiospores measuring 6.3-8.3 × 5.5-6.3 µm (Martínez & Nakasone, 2014) and 5-6.5(-7) × 4.5-6 µm (Parmasto, 1967, as H. caucasicum Parm.), and slightly thick-walled subicular hyphae. However, the two species have different substrate preferences, viz., N. asiaticum has been collected only from coniferous trees such as C. japonica and P. densiflora, whereas N. wakefieldiae have been recorded from both broad-leaved and coniferous trees (Nilsson & Hallenberg, 2003). Furthermore, ITS sequence data revealed that N. wakefieldiae is phylogenetically different from N. asiaticum, although sequences from the five specimens of the latter species do not form a single clade. At least N. asiaticum must currently be regarded as morphologically cryptic with N. wakefieldiae.
Neohypochnicium perlongicystidiosum N. Maek., Kogi & Norikura, sp. nov. (Figs. 3B, 5, 7B, 8B, 8D)
MycoBank No.: MB 842574.
Diagnosis: Neohypochnicium perlongicystidiosum is distinguished from others in the genus by having odontoid hymenophore, cylindrical and thick-walled cystidia (75-493 × 7.5-14.5 µm), and subglobose to globose, finely verrucose, thick-walled (up to 2.5 µm thick) basidiospores measuring (9-)10-12(-13.5) × (8.5-)9-11(-13) µm.
Holotype: TUMH 63328, on dead branch of a broad-leaved tree, Yakushima-cho (Yakushima Island), Kumage-gun, Kagoshima Pref., Japan, 6 Sep 2018, collected by N. Maekawa. Ex-holotype culture: TUFC 101573. GenBank: LSU = LC663690; ITS = LC663677.
Etymology: “perlongicystidiosum” refers to having significantly long cystidia.
Description: Basidiomes resupinate, adnate, effused; hymenial surface whitish cream, “Buff” to pale ochre, smooth, grandinioid to odontoid (5-18 warts per mm2), byssoid to hispidulous under the lens (×10); margin concolous to the hymenial surface, thinning out, indeterminate. In vertical section, subhymenium subhyaline, composed of densely interwoven hyphae; subiculum subhyaline, composed of loosely interwoven hyphae at first and then gradually becoming dense. Hyphal system monomitic; hyphae 4.5-8 µm wide, smooth, thin- to thick-walled (up to 2 µm thick), with clamp connection at all septa (Fig. 4D). Cystidia arising from subicular hyphae, cylindrical to tubular, 75-493 × 7.5-14.5 µm, smooth, thick-walled (2-4 µm thick) excepting the apical part, with a clamp at the basal septum, projecting up to 250 µm beyond the hymenial surface (Fig. 4C). Basidia clavate to suburniform, 46-70 × 9-12 µm, thick-walled (up to 1 µm thick) in the lower half, with a basal clamp, producing 4 sterigmata; sterigmata 7-9 µm in length (Fig. 4B). Basidiospores subglobose to globose, (9-)10-12(-13.5) × (8.5-)9-11(-13) µm (Lm = 11.2 ± 1.1 µm, Wm = 10.3 ± 1.0 µm, Q = 1.0-1.2, Qm = 1.1 ± 0.1; n=120/6), finely verrucose, thick-walled (up to 2.5 µm thick), cyanophilous, non-amyloid (Fig. 4A).
Characteristics in culture: The optimum growth temperature for the polysporous isolates TUFC 14478 and TUFC 101573 was 25 °C. Isolates could grow between 5 and 30 °C, but no visible growth was observed at 35 °C. Growth on MA was 25-28 mm in radius at 25 °C after 5 d in the dark. Mycelial mats at that time white, cottony to woolly (Fig. 8B); reverse side of mycelial mats not discolored after 2 w; agar bleached after 2 w; odor not noticeable. Aerial hyphae 2.5-8 µm wide, subhyaline, thin- to slightly thick-walled (up to 0.5 µm thick), clamp connections presnt at all septa, sometimes encrusted, moderately branched. Marginal hyphae 2.5-6.5 µm wide, subhyaline, thin-walled, clamp connections at all septa, sparsely branched. Submerged hyphae dense, 2.5-6.5 µm wide, subhyaline, sometimes with pale yellowish oily contents, thin- to slightly thick-walled (up to 0.5 µm thick), with clamp connections at all septa. Chlamydospores limoniform, fusiform to ellipsoid, sometimes irregular shaped, 27-52 × 14-25 µm, thick-walled (1.5-3 µm thick), terminal or intercalary, abundant on aerial hyphae (Fig. 8D). Conidia and cystidium-like cells not observed after 6 wk of incubation at 25 °C.
Species codes (Stalpers, 1978): 6, (12), 14, (21), (22), 30, 37, 39, 52, 53, 54, (55), (57), 82, 85.
Other specimens and cultures examined: JAPAN. TUMH 40397 on dead branch of a broad-leaved tree, Hachijo-machi (Hachijo Island), Tokyo, 8 Sep 2011, collected by N. Maekawa and R. Nakano (culture: TUFC 14478); TUMH 64602 on dead trunk of Quercus acutissima Carruth. (bedlog of shiitake cultivation), Hamatama-machi, Higashimatuura-gun, Saga Pref., 16 Dec 2003, collected by K. Kamohara; TUMH 64603 on dead trunk of Q. acutissima (bedlog of shiitake cultivation), Saigo-son, Higashiusuki-gun, Miyazaki Pref., 6 Dec 2003, collected by S. Komatsu; TUMH 64604 on dead trunk of Q. serrata Murray (bedlog of shiitake cultivation), Miyazaki Pref., 3 Feb 2016, collected by N. Maekawa; TUMH 63618 on dead trunk of Q. acutissima (bedlog of shiitake cultivation), Miyazaki Pref., 8 Jul 2016, collected by E. Nagasawa (culture TUFC 101628).
Remarks: Neohypochnicium perlongicystidiosum is primarily characterized by having grandinioid to odontoid hymenial surface and cylindrical, thick-walled cystidia measuring 75-493 × 7.5-14.5 µm. Neohypochnicium perlongicystidiosum is morphologically similar to H. longicystidiosum (S.S. Rattan) Hjortstam & Ryvarden (as Hyphodontia longicystidiosa S.S. Rattan; Rattan, 1977) and N. patagonicum in producing long cystidia. However, basidiospores of H. longicystidiosum are smooth and 4-5.5 × 4-4.5 µm (Rattan, 1977), whereas N. perlongicystidiosum produces finely verrucose and larger basidiospores measuring (9-)10-12(-13.5) × (8.5-)9-11(-13) µm. Neohypochnicium patagonicum produces also morphologically similar cystidia measuring 100-250(-350) × 8-12 µm, but its basidiospores [8-10 × (7-)7.5-8.5 µm, Gorjón & Hallenberg (2013)] are smaller than those of N. perlongicystidiosum. Furthermore, ITS sequence data showed that N. perlongicystidiosum is phylogenetically distinct from N. patagonicum.
Gyrophanopsis japonica N. Maek. & Kogi, sp. nov. (Figs. 3C, 6, 8C)
MycoBank No.: MB 842575.
Diagnosis: Gyrophanopsis japonica is characterized by byssoid to hispidulous hymenophore, cylindrical septocystidia (105-200 × 6-9 µm) heavily covered with subhyaline crystals, and broadly ellipsoid to subglobose, smooth and slightly thick-walled (up to 0.5 µm thick) basidiospores measuring 5-5.5 × 4-4.5 µm.
Holotype: TUMH 61400, on decayed wood of a broad-leaved tree, Taiwa-cho, Kurokawa-gun, Miyagi Pref., Japan, 21 Sep 2014, collected by N. Maekawa. Ex-holotype culture: TUFC 100860. GenBank: LSU = LC663688; ITS = LC663668.
Etymology: “japonica” refers to Japan, where the vouchered specimen was collected.
Description: Basidiome resupinate, adnate, effused, 100-200 µm thick; hymenial surface “Buff” when drying, smooth to grandinioid, byssoid to hispidulous under the lens (×10); margin concolorous with the hymenial surface, thinning out, indeterminate. In vertical section, subhymenium subhyaline, composed of densely interwoven hyphae; subiculum subhyaline, composed of loosely interwoven hyphae. Hyphal system monomitic; hyphae 4-9.5 µm wide, smooth, thin- to thick-walled (up to 2.5 µm thick), clamp connection at all septa, sometimes heavily encrusted. Cystidia (septocystidia) arising from subicular hyphae, cylindrical with obtuse apex, 2-4 clamped septate, 105-200 × 6-9 µm, thick-walled (up to 2 µm thick), but wall becoming thinner towards the apex, with a clamp connection at the basal septum, projecting up to 130 µm beyond the hymenial surface, heavily encrusted with subhyaline crystalline material of 1-2 µm thick. Basidia subclavate to subcylindric, slightly constricted, 14-20 × 5-5.5 µm, with a basal clamp, producing 4 sterigmata; sterigmata 2-3 µm in length. Basidiospores broadly ellipsoid to subglobose, 5-5.5 × 4-4.5 µm [Lm = 5.4 ± 0.2 µm, Wm = 4.2 ± 0.3 µm, Q = 1.2-1.4, Qm = 1.3 ± 0.1; n=20/1], smooth, slightly thick-walled (up to 0.5 µm thick), cyanophilous, non-amyloid.
Characteristics in culture: The optimum growth temperature for the polysporous isolate TUFC 100860 was 25-30 °C. Isolate could grow between 5 and 35 °C, but no visible growth was observed at 40 °C. Growth on MA was 22-28 mm in radius at 25 °C after 21 d in the dark. Mycelial mats at that time transparent, but whitish around the inoculum, downy; reverse side of mycelial mats not discolored; agar not bleached. Aerial hyphae 2-4 µm wide, subhyaline, thin-walled, usually lacking clamp connections at the septa, moderately branched, occasionally encrusted. Marginal hyphae 2-4 µm wide, subhyaline, thin-walled, lacking clamp connections at all septa, sparsely branched. Submerged hyphae 2-4.5 µm wide, subhyaline, thin-walled, clamp connections at the septa, but occasionally clampless septate, moderately branched. Chlamydospores, conidia and cystidium-like cells not observed after 3 w of incubation at 25 °C.
Species codes (Stalpers, 1978): 9, 13, 14, 17, (30), (39), 52, 53, (82).
Specimen and culture examined: see holotype.
Remarks: Gyrophanopsis japonica is characterized by its wider subicular hyphae (4-9.5 µm wide), heavily encrusted septocystidia, small basidia (14-20 × 5-5.5 µm) and globose, slightly thick-walled basidiospores measuring 5-5.5 × 4-4.5 µm. This species differs phylogenetically from the previously described two species, Gy. polonensis and G. zealandica. In addition, Gy. japonica produces globose basidiospores (5-5.5 × 4-4.5 µm) and basidia measuring 14-20 × 5-5.5 µm, whereas Gy. polonensis produces ellipsoid basidiospores (6.5-8.5 × 4-5 µm) and longer basidia measuring 20-35 × 4-6 µm (Maekawa, 1994). On the other hand, G. zealandica strongly resembles Gy. japonica, but differs in some minor characters. According to Stalpers and Buchanan (1991), septocystidia of G. zealandica are partly covered with a sheath of yellowish-brown crystalline material of about 1 µm thick, whereas those of Gy. japonica are covered with subhyaline crystalline material. The other morphological characteristics, viz. basidiospore shape and size and hyphal features of Gy. japonica are overlapped to those of Gy. zealandica.

Fig. 3 - Hymenial surface of basidiomata. A: Neohypochnicium asiaticum (TUMH 61220, holotype). B: N. perlongicystidiosum (TUMH 63328, holotype). C: Gyrophanopsis japonica (TUMH 61400, holotype). Bars: 1 mm.

Fig. 4 - Microscopic elements of Neohypochnicium asiaticum (TUMH 61220, holotype). A: Basidiospores. B: Basidia. C: Subicular hyphae. D: Cystidia. Bars: 10 µm.

Fig. 5 - Microscopic elements of Neohypochnicium perlongicystidiosum (TUMH 63328, holotype). A: Basidiospores. B: Basidia. C: Subicular hyphae. D: Cystidia. Bars: A-C 10 µm; D 20 µm.
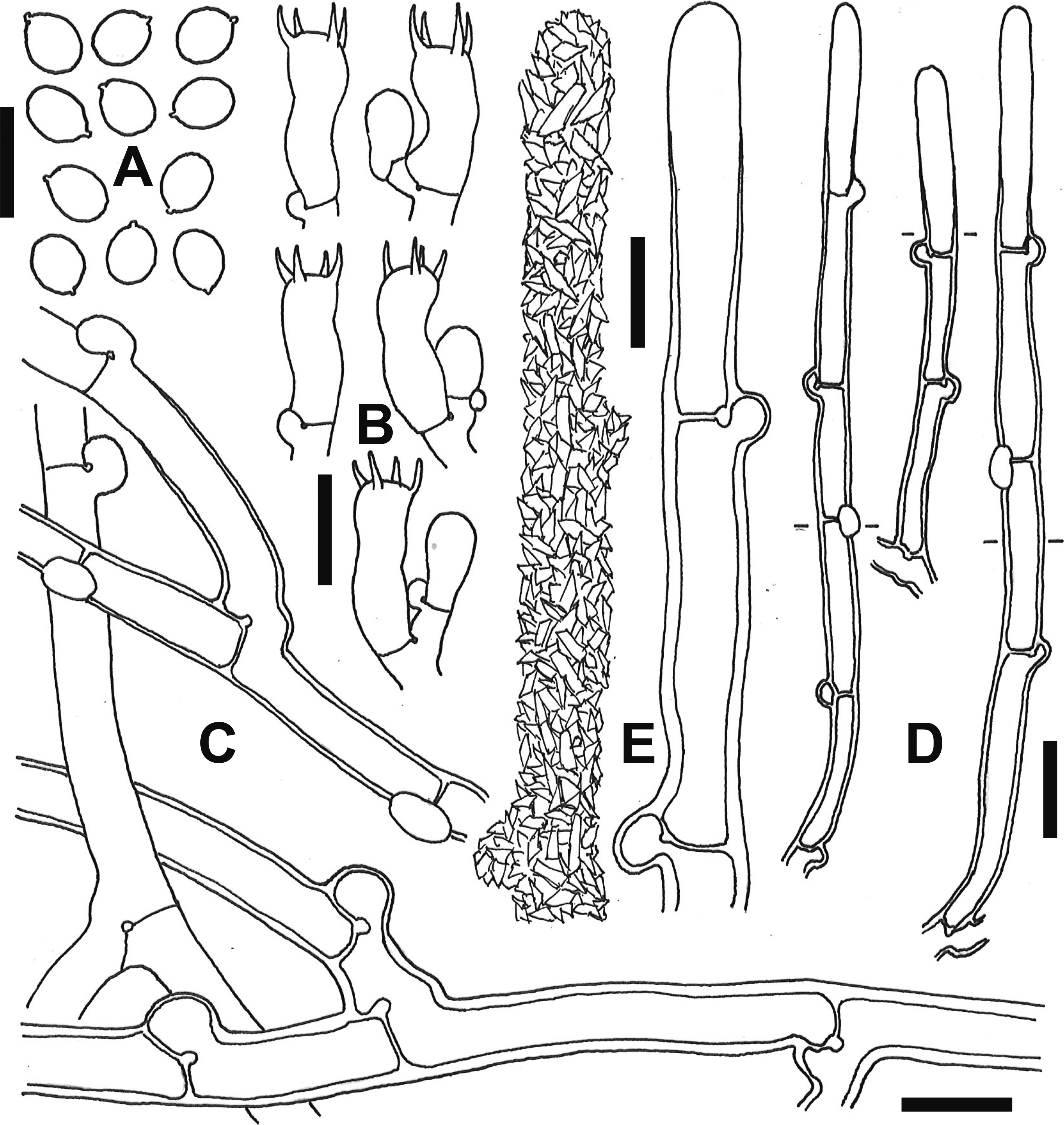
Fig. 6 - Microscopic elements of Gyrophanopsis japonica (TUMH 61400, holotype). A: Basidiospores. B: Basidia. C: Subicular hyphae. D: Cystidia. E: Surface (left) and outline (right) of upper part of a cystidium. Bars: A-C, E 10 µm; D 20 µm.

Fig. 7 - Scanning electron photographs of basidiospores. A: Neohypochnicium asiaticum (TUMH 61220, holotype). B: N. perlongicystidiosum (TUMH 63328, holotype). C: N. subrigescens (TUMH 64612). Bars: 10 µm.

Fig. 8 - Mycelia and chlamydospores produced in culture. A: Neohypochnicium asiaticum (ex-holotype culture: TUFC 31698) on MA at 25 °C after 5-d incubation. B: N. perlongicystidiosum (ex-holotype culture: TUFC 101573) on MA at 25 °C after 5-d incubation. C: Gyrophanopsis japonica (ex-holotype culture: TUFC 100860) on MA at 25 °C after 20-d incubation. D: Chlamydospores of N. perlongicystidiosum (TUFC 101573) produced on MA after 7-d incubation at 25 °C. Bar: 50 µm.
Neohypochnicium albostramineum (Bres.) N. Maek., comb. nov.
Basionym: Hypochnus albostramineus Bres., Annls mycol. 1(2): 109, 1903.
MycoBank No.: MB 842576.
Remarks: For descriptions, see Bernicchia and Gorjón (2010), and Nilsson and Hallenberg (2003). This species has broadly ellipsoid, finely verrucose basidiospores measureing 8-10(-12) × 6.5-7(-8) µm (Bernicchia & Gorjón, 2010).
Neohypochnicium aotearoae (B.C. Paulus, H. Nilsson & Hallenb.) N. Maek., comb. nov.
Basionym: Hypochnicium aotearoae B.C. Paulus, H. Nilsson & Hallenb., N.Z. Jl Bot. 45(1): 144, 2007.
MycoBank No.: MB 842577.
Remarks: This species has ovoid to subglobose, verrucose basidiospores measuring 5.6-8.7 × 4.8-7.3 µm (Paulus, Nilsson & Hallenberg, 2007).
Neohypochnicium capitulatum (Boidin & Gilles) N. Maek., comb. nov.
Basionym: Hypochnicium capitulatum Boidin & Gilles, Bull. Soc. Mycol. Fr., 116: 164, 2000.
MycoBank No.: MB 842578.
Remarks: Although sequence data of N. capitulatum has not been obtained, the species belongs to Neohypochnicium because it forms subglobose, warted basidiospores measuring 8-9 × 7-7.5 µm (Boidin & Gilles, 2000).
Neohypochnicium cremicolor (Bres.) N. Maek., comb. nov.
Basionym: Hypochnus cremicolor Bres., Annls mycol. 1(2): 109, 1903.
MycoBank No.: MB 842579.
Remarks: For descriptions, see Bernicchia and Gorjón (2010), and Nilsson and Hallenberg (2003). This species has broadly ellipsoid, finely verrucose basidiospores measuring 6-6.5 × 5-5.5 µm (Bernicchia & Gorjón, 2010).
Neohypochnicium cystidiatum (Boidin & Gilles) N. Maek., comb. nov.
Basionym: Hypochnicium cystidiatum Boidin & Gilles, Cahiers de La Maboké, 9(2): 90, 1971.
MycoBank No.: MB 842580.
Remarks: This species has ovoid, ornamented basidiospores measuring (4.8-)5.2-6.7 × 4.2-5.2 µm (Boidin & Lanquetin, 1971).
Neohypochnicium geogenium (Bres.) N. Maek., comb. nov.
Basionym: Corticium geogenium Bres., Annls Mycol. 1(2): 98, 1903.
MycoBank No.: MB 842581.
Remarks: For descriptions, see Bernicchia and Gorjón (2010), Eriksson and Ryvarden (1976). This species has ellipsoid, smooth basidiospores measuring 6-7.5(-9) × 4-4.5(-5.5) µm (Eriksson & Ryvarden, 1976).
Neohypochnicium guineense (Tellería, M. Dueñas, Melo & M.P. Martín) N. Maek., comb. nov.
Basionym: Hypochnicium guineense Tellería, M. Dueñas, Melo & M.P. Martín, Mycologia 102(6): 1431, 2010.
MycoBank No.: MB 842582.
Remarks: This species has globose, verrucose basidiospores measuring (7-)8-9 × (7-)7.5-8.5(-10) µm (Tellería et al., 2010).
Neohypochnicium huinayense (Tellería, M. Dueñas & M.P. Martín) N. Maek., comb. nov.
Basionym: Hypochnicium huinayense Tellería, M. Bueñas & M.P. Martín, Persoonia 31: 281, 2013.
MycoBank No.: MB 842583.
Remarks: This species has globose, ornamented basidiospores measuring 6.5-8(-9) × 6.5-8 µm (Crous et al., 2013).
Neohypochnicium michelii (Tellería, M. Dueñas, Melo & M.P. Martín) N. Maek., comb. nov.
Basionym: Hypochnicium michelii Tellería, M. Dueñas, Melo & M.P. Martín, Mycologia 102: 1431, 2010.
MycoBank No.: 842585.
Remarks: This species has ellipsoid, smooth basidiospores measuring (7.5-)9-11 × (6-)7-7.5 µm (Tellería et al., 2010).
Neohypochnicium microsporum (G. Gruhn, Schimann & M. Roy) N. Maek., comb. nov.
Basionym: Hypochnicium microsporum G. Gruhn, Schimann & M. Roy, Bull. Soc. Mycol. Fr., 130: 303, 2017.
MycoBank No.: MB 842584.
Remarks: This species has subglobose, smooth basidiospores measuring (3.7-)3.9-5(-5.8 × (3.2-)3.8-5(-5.3) µm (Gruhn, Schimann & Roy, 2014).
Neohypochnicium patagonicum (Gorjón & Hallenb.) N. Maek., comb. nov.
Basionym: Hypochnicium patagonicum Gorjón & Hallenb., Mycol. Progr. 12(2): 188, 2013.
MycoBank No.: MB 842586.
Remarks: This species has broadly ellipsoid to subglobose, verrucose basidiospores measuring 8-10 × (7-)7.5-8.5 µm (Gorjón & Hallenberg, 2013).
Neohypochnicium pini (Y. Jang & J-J. Kim) N. Maek., comb. nov.
Basionym: Hypochnicium pini Y. Jang & J-J. Kim, Mycotaxon 124: 211, 2013.
MycoBank No.: MB 842587.
Description: Basidiomes resupinate, adnate, effused, 50-200 µm thick; hymenial surface white, “Buff” to “Pale Luteous” when drying, smooth to minutely warted. Hyphae 4-6 µm wide, smooth, thin- to thick-walled (up to 2 µm thick), with clamp connection at all septa. Cystidia cylindrical to subfusiform with obtuse apex, 53-132 × 6-11.5 µm, thin- to slightly thick-walled (up to 0.5 µm), with a clamp connection at the basal septum, projecting up to 50 µm beyond the hymenial surface. Basidia clavate to suburniform, 18-37 × 5-8 µm, producing 4 sterigmata. Basidiospores broadly ellipsoid, 5.5-7(-8) × 4.5-6(-6.5) µm, finely verrucose, thick-walled (up to 1 µm thick), cyanophilous, non-amyloid.
Specimens and cultures examined: JAPAN. TUMH 64589 [as H. punctulatum (TMI 6991)] on dead trunk of a broad-leaved tree, Daisen-cho, Saihaku-gun, Tottori Pref., 18 Oct 1981, collected by N. Maekawa; TUMH 64588 [as H. punctulatum (TMI 6771)] on dead trunk of Pinus densiflora, Ichikawa-cho, Kanzaki-gun, Hyogo Pref., 19 Jul 1981, collected by N. Maekawa; TUMH 64590 [as H. punctulatum (TMI 7618)] on dead trunk of Quercus serrata, Higashioro, Tottori City, Tottori Pref., 9 Oct 1983, collected by N. Maekawa; TUMH 64592 [as H. punctulatum (TMI 17547)] on decayed wood of a broad-leaved tree, Daisen-cho, Saihaku-gun, Tottori Pref., 1 Oct 1984, collected by N. Maekawa; TUMH 61221 [as H. punctulatum (TMI 17546)] on dead trunk of Castanopsis cuspidata (Thunb.) Schottky, Ue-machi, Tottori City, Tottori Pref., 1 Jul 1985, collected by N. Maekawa and E. Nagasawa (culture: TUFC 30475); TUMH 64591 on dead branch of Larix kaempferi (Lamb.) Carrière, Miasa-mura, Kitaazumi-gun, Nagano Pref., 1 Oct 1987, collected by N. Maekawa; TUMH 64587 on dead branch of a broad-leaved tree, Yorii-machi, Osato-gun, Saitama Pref., 12 Sep 2014, collected by H. Kogi.
Remarks: Of the Japanese specimens, TUMH 64588, TUMH 64589, TUMH 64590, TUMH 64592 and TUMH 61221 were reported as H. punctulatum by Maekawa (1994), but in this study these specimens were identified as N. pini based on phylogenetic analyses and morphological re-observation.
Neohypochnicium punctulatum (Cooke) N. Maek., comb. nov.
Basionym: Corticium punctulatum Cooke, Grevillea 6(no. 40): 132, 1878.
MycoBank No.: MB 842588.
Description: Basidiomes resupinate, adnate, effused, 70-110 µm thick; hymenial surface white, “Buff” to “Pale Luteous” when drying, smooth. Hyphae 4-6 µm wide, smooth, thin-walled, with clamp connection at all septa. Cystidia cylindrical to subfusiform with obtuse apex, 70-107 × 5-10 µm, thin-walled, with a clamp connection at the basal septum, projecting up to 30 µm beyond the hymenial surface. Basidia clavate to suburniform, 28-35 × 6.5-7.5 µm, with a basal clamp, producing 4 sterigmata. Basidiospores broadly ellipsoid, 7.5-8.5 × 6.5-7.5 µm, finely verrucose, thick-walled (up to 1 µm thick), cyanophilous, non-amyloid.
Specimen and culture examined: JAPAN. TUMH 61188 on dead trunk of dead trunk of a coniferous tree, Iijima-machi, Kamiina-gun, Nagano Pref., 10 Oct 1996, collected by N. Maekawa (culture: TUFC 33598).
Remarks: Basidiospore dimensions of the Japanese specimen (TUMH 61188) fall into those of H. punctulatum s. str. described by Nilsson and Hallenberg (2003). The Japanese specimens reported as H. punctulatum and H. eichleri by Maekawa (1994) were identified N. asiaticum (see specimens of N. asiaticum) or N. pini (see specimens of N. pini) based on the phylogenetic analyses and morphological re-observation.
Neohypochnicium subrigescens (Boidin) N. Maek., comb. nov.
Basinonym: Hypochnicium subrigescens Boidin, Cahirs de La Maboké, 9(2): 90, 1971.
MycoBank No.: MB 842589.
Description: Basidiomes resupinate, adnate, effused, 100-200 µm thick; hymenial surface white to “Buff” when drying, smooth. Hyphae 3-8 µm wide, smooth, thin- to slightly thick-walled (up to 0.5 µm thick), with clamp connections at all septa, densely arranged in the subiculum. Cystidia cylindrical to subfusiform with obtuse apex, 42-180 × 6.5-10 µm, thin-walled, with a clamp connection at the basal septum, projecting up to 100 µm beyond the hymenial surface. Basidia clavate to suburniform, 18-36 × 5-8 µm, with a basal clamp, producing 4 sterigmata. Basidiospores subglobose to broadly ellipsoid, (5.5-)6-7(-7.5) × (5-)5.5-6.5(-7) µm, smooth, thick-walled (up to 1 µm thick), weakly cyanophilous, non-amyloid.
Specimens examined: JAPAN: TUMH 61539 on decayed wood of Cerasus sargentii (Rehder) H. Ohba, near Hanadate, Osaki City, Miyagi Pref., 22 Sep 2014, collected by N. Maekawa (culture: TUFC 100892); TUMH 64583 [as H. sphaerosporum (TMI 10702)] and TUMH 64584 [as H. sphaerosporum (TMI 17551)] on a shiitake bedlog of Quercus serrata, Daito-cho, Higashiiwai-gun, Iwate Pref., 5 Nov 1986, collected by N. Maekawa; TUMH 64585 [as H. sphaerosporum (TMI 17552)] on a shiitake bedlog of Q. crispula Blume, Daito-cho, Higashiiwai-gun, Iwate Pref., 5 Nov 1986, collected by N. Maekawa; TUMH 64586 [as H. sphaerosporum (TMI 17553)] on decayed wood of a broad-leaved tree, Misakubo-cho, Iwate-gun, Iwate Pref., 21 Sep 1986, collected by N. Maekawa, collected by N. Maekawa; TUMH 64582 [as H. sphaerosporum (TMI 9477)] on dead trunk of Pinus sp., Miasa-mura, Kitaazumi-gun, Nagano Pref., 1 Oct 1987, collected by N. Maekawa; TUMH 64612 on dead and decorticated trunk of a coniferous tree, Takane-machi, Takayama City, Gifu Pref., 27 Sep, 2015, collected by N. Maekawa. DENMARK: GB-0150192 on log of Betula sp., Jutland, Ebeltoft, Ahl, 1 Sep 1987, collected by N. Hallenberg. NORWAY: GB-0104172 on dead Alnus incana (L.) Moench, Akershus, Skedsmo, Holmen Nature Reserve, 1994, collected by H. Kauserud. SWEDEN: GB-0087503, Västergötland, Medelplana par., Rabäck, Munkängarna, 8 Oct 2008, collected by K.H. Larsson; GB-0090069 on deciduous wood, Västergötland, Nolhaga, 19 Sep 1994, collected by K. Hjortstam; GB-0090501 on wood of angiosperm, Västergötland. V:a Tunhem par., Halleberg, Hallesnipen-Ovandalen, 5 Oct 2006, collected by K.H. Larsson; GB-0090583 on Betula sp., Jämtland, Kall par., Skäckerfjällen Nature Res., 6 Sep 2006, collected by K.H. Larsson.
Remarks: This species is characterized by densely arranged relatively thin-walled hyphae in the subiculum and smooth, subglobose to broadly ellipsoid basidiospores measuring (5.5-)6-7(-7.5) × (5-)5.5-6.5(-7) µm (Fig. 7C). Hypochnicium erikssonii Hallenb. & Hjortstam resembles H. subrigescens morphologically, but differs in having larger basidiospores measuring 7-8 × 5.5-7 µm (Hallenberg & Hjortstam, 1990). In Japan, Maekawa (1994) reported a Hypochnicium species with smooth, subglobose to globose basidiospores measuring 6-7.5 × 5-6.5 µm as H. sphaerosporum. But H. sphaerosporum has been treated a synonym of H. punctulatum because its type specimen produces weakly ornamented basidiospores (Hallenberg & Hjortstam, 1990; Nilsson & Hallenberg, 2003). In the present study, reexamination of the Japanese specimens of H. sphaerosporum (TUMH 64582, TUMH 64583, TUMH 64584, TUMH 64585 and TUMH 64586) showed their smooth basidiospores. In addition, the morphological features of these Japanese specimens were identical to those of the six specimens of H. subrigescens collected from Denmark, Norway and Sweden.
Neohypochnicium wakefieldiae (Bres.) N. Maek., comb. nov.
Basionym: Corticium wakefieldiae Bres., Annls mycol., 18(1/3): 48, 1920.
MycoBank No.: MB 842590.
Remarks: For descriptions, see Bernicchia and Gorjón (2010), Nilsson and Hallenberg (2003), and Tellería et al. (2010). This species has broadly ellipsoid, finely verrucose basidiospores measureing 6.5-8 × 5.5-6 µm (Bernicchia & Gorjón, 2010).
3.2.4. Previously unreported species from Japan
Hypochnicium multiforme (Berk. & Broome) Hjortstam, Windahlia 23: 2, 1998.
Description: Basidiomes resupinate, adnate, effused, 50-150 µm thick; hymenial surface grayish white to pale yellowish white when drying, smooth; margin concolous with the hymenial surface, thinning out, indeterminate. Hyphae 2-5(-6) µm wide, smooth, thin- to slightly thick-walled (up to 1 µm thick), with clamp connections at all septa, densely arranged in the subiculum. Cystidia lacking. Basidia subclavate to suburniform, 22-45 × 7.5-12 µm, with a basal clamp, producing 4 sterigmata. Basidiospores subglobose to globose, (7-)9-11(-12) × (6.5-)8-10(-11) µm, smooth, thick-walled (up to 2 µm thick), weakly cyanophilous, non-amyloid.
Specimens and cultures examined: JAPAN. TUMH 64580 [as H. globosum (TMI 10705)] on decayed wood of Pinus densiflora, Ue-machi, Tottori City, Tottori Pref., 10 Jul 1986, collected by N. Maekawa; TUMH 64579 [as H. globosum (TMI 15329)] on decayed wood of P. densiflora, Nishigo-mura, Nishishirakawa-gun, Fukushima Pref., 2 Nov 1986, collected by N. Maekawa; TUMH 64578 [as H. globosum (TMI 15330)] on dead branch of a broad-leaved tree, Aihara, Sagamihara City, Kanagawa Pref., 6 Oct 1991, collected by S. Kigawa; TUMH 61219 on dead trunk of Trochodendron aralioides Siebold & Zucc., Kumage-gun (Yakushima Island), Kagoshima Pref., 25 Oct 1995, collected by N. Maekawa; TUMH 40197 on dead branch of a broad-leaved tree, Midorigaoka, Iwamizawa City, Hokkaido, 25 Sep 2010, collected by N. Maekawa (culture: TUFC 12763); TUMH 64581 on dead branch of Quercus crispula Blume, Takane-machi, Takayama City, Gifu Pref., 28 Sep 2015, collected by N. Maekawa.
Remarks: The Japanese specimens are primarily characterized by having ovoid to subglobose, smooth, thick-walled (up to 2 µm thick) and weakly cyanophilous basidiospores measuring (7-)9-11(-11.5) × (6.5-)8-10(-11) µm, and lacking cystidia. These characteristics conform the descriptions of H. karstenii (Bres.) Hallenb. (Hallenberg, 1983), a synonym of H. multiforme.