Abstract
ダイズとラッカセイ以外の食用マメ科作物は,雑豆(ざっとう)と呼ばれる.雑豆はゲノム構造が単純な種を含み,かつ種多様性に富むことから,研究材料という点でダイズより優れているが,ダイズの育種素材(遺伝資源)として直接利用することはできない.しかし,雑豆から有用遺伝子の情報を引き出すことができれば,類似した遺伝的背景を保有するダイズの育種に応用できるはずである.本稿では,栽培化形質のうち裂莢性の消失を例に,ダイズ育種における雑豆の遺伝子情報の利用可能性を概説したい.
はじめに
雑豆(ざっとう)とは,ダイズ(Glycine max (L.) Merr.)とラッカセイ(Arachis hypogaea L.)以外の食用マメ科作物の総称で,我が国ではアズキ(Vigna angularis (Willd.) Ohwi et H. Ohashi),インゲンマメ(Phaseolus vulgaris L.),エンドウマメ(Pisum sativum L.),ソラマメ(Vicia faba L.)が良く知られている.
実のところ,研究材料という観点では,雑豆はダイズより優れている点が多い.例えば,ダイズは古倍数性の 2 倍体でほとんどの遺伝子が重複している上,ゲノムサイズが約 11 億塩基対もあるが,アズキは一般的な 2 倍体でゲノムサイズが約 5 億塩基対と小さいため遺伝解析が容易である(Sakai et al. 2015).また,同属内の栽培化過程の比較遺伝解析の面においても,雑豆はダイズより優れている.Glycine 属で栽培化された種はダイズのみであるが,Vigna 属ではアズキ,リョクトウ(Vigna radiata (L.) Wilczek),ササゲ(Vigna unguiculata (L.) Walp.)を含む約 10 種が独立に栽培化されている(Kaga et al. 2011).そのため Vigna 属の栽培種間では,類似した遺伝的背景において,それぞれの種が独立して獲得した多様な栽培化遺伝子の比較解析が可能である.
作物は自然に生息する野生植物から約 1 万年から数千年かけて現在の姿となった.農耕を開始した人類が野生植物を採集して,栽培と収穫を繰り返す中で,意識的または無意識に自然突然変異による遺伝形質を選抜したのだ.この過程を栽培化,選抜された遺伝形質を栽培化形質,栽培化形質の責任遺伝子を栽培化遺伝子と呼ぶ.食用マメ科作物における主要な栽培化形質は,裂莢性の消失,硬実性の消失,種子の大型化である(Abbo et al. 2014,Fuller et al. 2014,Smýkal et al. 2014,Fuller and Allaby 2009).
ダイズと雑豆の栽培化形質を比較してみると,同一の形質が異なるメカニズムで生み出されてきたことが分かる.例えば,ダイズの種子は種皮全体が透水性になり,硬実性が消失したと考えられるが(Koizumi et al. 2008),アズキやインゲンマメの種子はレンズと呼ばれる組織のみが透水性になっている(Kikuchi et al. 2006).このため,ダイズの種子浸水直後の吸水速度は雑豆よりも早く(上中ら 2000),雑豆よりもダイズの硬実性の消失程度は大きい.一方,以降に概説するように,裂莢性の消失については,ダイズより雑豆のほうが進んでいる.
残念なことに,ダイズと雑豆間での属間交雑の成功例はこれまで報告されておらず,雑豆をダイズ育種の素材として直接利用することはできない.しかし,雑豆から多様な栽培化遺伝子の情報を引き出せば,ダイズの育種に応用できるはずだ.
裂莢性の消失
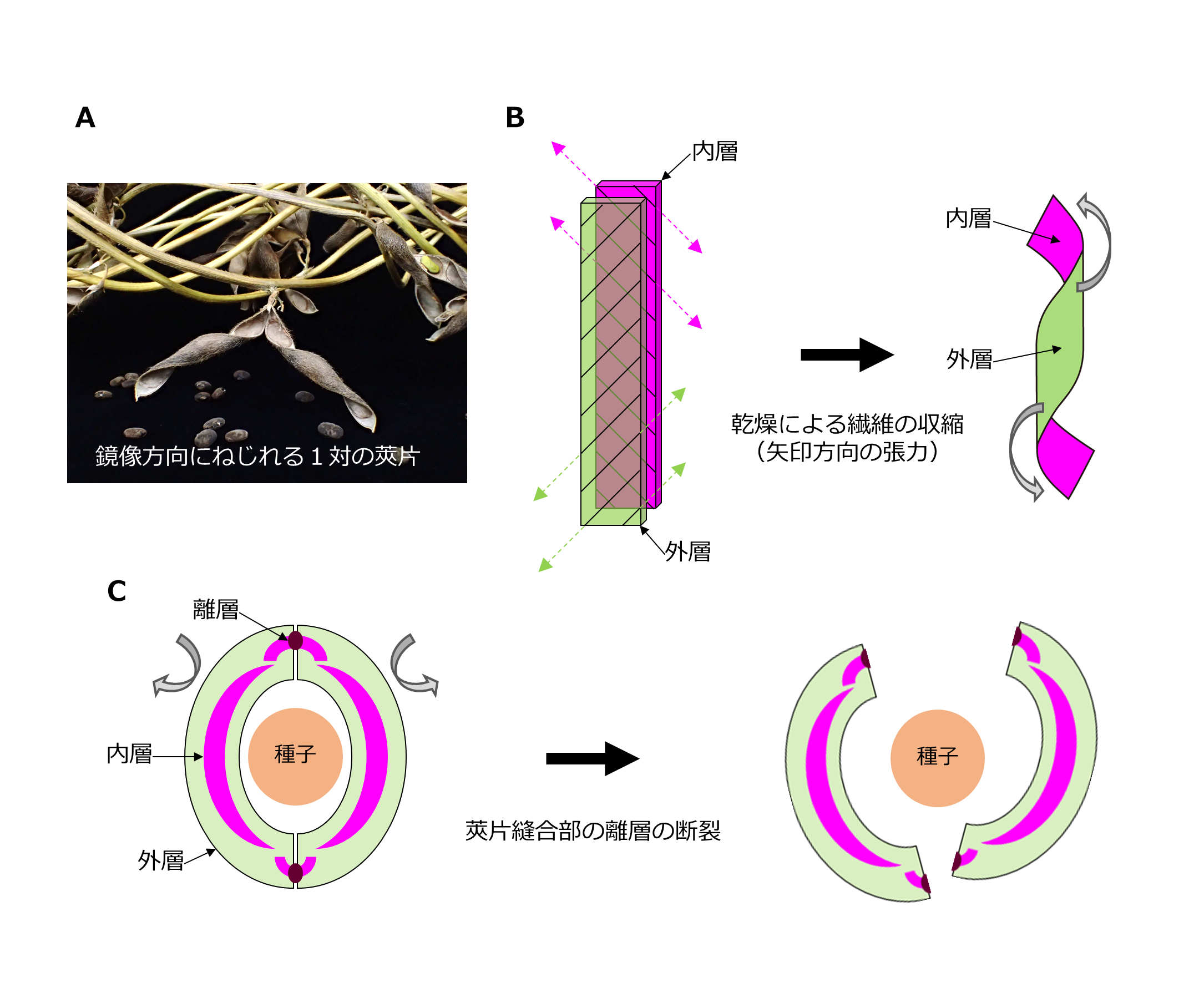
自然環境においてマメ科植物の莢は,成熟後の乾燥により裂莢し,種子を飛散させる(図 1A).マメ科植物の裂莢には「莢片のねじれ力」と「莢片縫合部の離層形成」が重要である.莢片に発達する外層と内層の繊維は,互いに 45 度ずれていて,乾燥につれて垂直方向に収縮することで,ねじれ力を生じる(図 1B)(Erb et al. 2013, Armon et al. 2011).1 対の莢片は,お互い鏡像方向にねじれるため,莢片縫合部の離層にひずみが生じて断裂すると,抑制されていたねじれ力が解放されて裂莢する(図 1C).この力は平地において種子を 7 m も飛散させることがあり(Yoshimura et al. 2011),動物と違って移動することができない植物が,生息域を拡大するための原動力となっている.
約 1 万年にもわたる人為選抜にも関わらず,ダイズの裂莢は収量を低下させる問題として残されている.湿潤気候の日本では,ダイズ品種「サチユタカ」をコンバイン収穫した際の全裂莢損失率は最大 63%に達することがあり(Yamada et al. 2017),乾燥気候におけるダイズの収量損失率は最大 100%になることもある(Bhor et al. 2014).裂莢の難易には品種間差があり,裂莢に至る相対湿度の評価では,ダイズ品種「Rokusun」「Ogden」「Lee」は,それぞれ相対湿度が 60%,30%,15%で裂莢した(Caviness 1965).気候変動により世界の平均湿度が下降すると予測されており(Sherwood and Fu 2014),今後数十年間の乾燥化で裂莢の問題が深刻化すると考えられている(Parker et al. 2021).このため,乾燥状態においても裂莢が起こりにくい形質 (難裂莢性)を持つダイズ品種の開発が急務である.
このような遺伝子型と環境の相互作用により生じる裂莢が招く収量損失を最小限にするためには,「莢片のねじれ力」と「莢片縫合部の離層形成」に関与する責任遺伝子とその環境応答性を解明することが重要である.ダイズの裂莢に関与する主要な量的形質遺伝子座(QTL)が第 16 染色体にマッピングされた後(Funatsuki et al. 2006),ダイズの難裂莢品種は Pod dehiscence 1(Pdh1)遺伝子の途中に終始コドンを生じていることが明らかとなった(Funatsuki et al. 2014).この遺伝子の変異は,既知の顕微的形質と関連していないが,莢片のねじれ力を減少させることから,莢片の木質層の配向など質的な影響を与えている可能性が高い(図 2A, B, E, H).SHATTERING1-5(SHAT1-5)遺伝子の機能獲得型変異も,ダイズの裂莢の低減に関連している (Dong et al. 2014).遺伝子上流のリプレッサーエレメントに生じた 20bp の欠失変異は SHAT1-5 の遺伝子発現を増加させ,離層付近の木質を厚くする(図 2A, B, E, H).しかし,多くの栽培品種では,依然として野生型対立遺伝子が優勢であり,この遺伝子座の効果は Pdh1 と比較して小さい(Zhang and Singh 2020).Pdh1 遺伝子の発見により日本のダイズ品種の収量損失は飛躍的に低下したが,難裂莢型 pdh1 対立遺伝子をもつ「サチユタカ A1」でも裂莢損失率は最大で約 20%に達することから,Pdh1 遺伝子座のみでは裂莢の問題を完全に克服できないことも分かった(Yamada et al. 2017).同じように,米国ではほぼ全ての品種が難裂莢型 pdh1 対立遺伝子を有しているが未だに裂莢が問題となっており,世界的な裂莢の問題解決には新たな遺伝変異の利用が欠かせない.最近になって,SHAT1-5 のパラログである NST1A の機能獲得型対立遺伝子がダイズの難裂莢性に寄与する可能性が報告された(Zhang and Singh 2020).難裂莢性を対象としたゲノムワイドアソシエーション解析によって検出された ss.715598106 SNP は,NST1A に終始コドンを生じる ss.98955957 SNP から 49 kb も離れているが,SHAT1-5 の SNP よりも難裂莢性との関連が高いため,遺伝子機能の証明を含む追加解析を行ったうえで,利用を検討する価値があろう.

雑豆にはダイズとは異なる難裂莢形質がみられ,最近になってその責任遺伝子が同定され始めている.アズキにおける裂莢は,莢片の木質層の消失と関連している(図 2A, C, F, I).莢片のねじれ数に基づく QTL 解析によって,第 7 染色体に存在する単一の潜性遺伝子が裂莢性の消失を支配していることが報告された後(Kaga et al. 2008,Isemura et al. 2007),戻し交配集団を用いたファインマッピングにより QTL が 4 kbp に絞り込まれた(Takahashi et al. 2020).この領域には,VaMYB26(Vigan.07g034400)のみがアノテーションされている.VaMYB26は,インゲンマメやササゲの難裂莢性の責任遺伝子の候補である PvMYB26(Phvul.005G157600)や VuMYB26(Vigun05g273500)のオルソログである(Di Vittori et al. 2021,Parker et al. 2020,Watcharatpong et al. 2020,Rau et al. 2019,Lo et al. 2018).野生アズキと栽培アズキの VaMYB26 の配列比較により,栽培アズキのタンパク質翻訳領域にチミンが挿入され,フレームシフトを生じることが判明している.VaMYB26 の変異は,栽培アズキで木質層の消失または減少を引き起こすことで,莢片のねじれ力を激減させる.アズキの VaMYB26 の変異は,ダイズの Pdh1 の変異よりも遥かに莢片のねじれ力を抑制するため,ダイズの難裂莢を向上させる育種において利用すべき遺伝子の変異の一つである.雑豆では,莢片のねじれは木質層の厚さと強い正の相関があることが知られている.ダイズやマメ科野生種は厚い木質層の莢片を持っており,この厚い木質層は裂莢を促進するだけでなく,光合成産物を種子と競合して収量を損なうと考えられている(Takahashi et al. 2020,Murgia et al. 2017,Assefa et al. 2013).そのため,木質層を消失させる VaMYB26 のような変異はダイズの子実収量の向上に貢献する可能性もある.
一方,著者の知る限り「莢片縫合部の離層形成」の抑制に関して最も強い影響を与える変異は,南インドで半栽培されているハネアズキ(Vigna stipulacea Kuntze)の変異原処理集団から選抜された変異体 rps1 に見られる(Takahashi et al. 2019).この変異体 rps1 は,莢片縫合部の離層が完全に消失しており,ダイズの SHAT1-5 の変異よりも遥かに高い裂莢抑制効果を示す(図 2A, D, G, J).顕微的観察において,変異体 rps1 は,ルピナス属植物の自然変異から Gladstones (1967)が選抜した変異体(invulnerabilis, tardus, conjunctus)と酷似している(Parker et al. 2021).既に,変異体 rps1 の責任遺伝子は,潜性の単一遺伝子であることが明らかとなっており(データ未公開),今後の機能解析が待たれる.
雑豆に学ぶダイズの育種
高橋,友岡(2017)は,雑豆の栽培化遺伝子の情報を活用したダイズの育種を提案している.上述のように,アズキの VaMYB26 は「莢片のねじれ力」に関して,ハネアズキ rps1 変異体の責任遺伝子は「莢片縫合部の離層形成」に関して,それぞれ潜性の単一遺伝子座により,ダイズの Pdh1 や SHAT1-5 の変異よりも裂莢の抑制に高い効果を示す(私信).したがって,突然変異育種により,これら遺伝子の変異を誘発することで,裂莢が完全に抑制されたダイズを開発できる可能性がある.
ただし,ダイズのようなパラロガス遺伝子対が部分的に保持される古倍数体に突然変異育種を適用する際には,非倍数性の 2 倍体作物より効率が悪いことを考慮する必要がある.ダイズと雑豆の共通祖先は 5,900 万年前に全ゲノム重複を経験した後,ダイズの祖先は約 1,300 万年前に 2 度目の全ゲノム重複を経験した(Schmutz et al. 2010).このため,2 倍体の雑豆が 1 組の遺伝子を持つ場合,ダイズではその遺伝子をパラロガス遺伝子対として 2 組保有することが多い(Wang et al. 2017).単一遺伝子の機能欠損に起因する雑豆の形質変異をダイズで利用するには,ダイズにおけるパラロガス遺伝子対の両方を機能欠損させる必要がある.
つまり,ダイズに雑豆の優れた難裂莢形質を付与するには,その責任遺伝子の各パラログの機能欠損変異体を変異原処理集団から逆遺伝的にスクリーニングし,その後,交雑により二重変異体を育成する手法が有効である.逆遺伝学的選抜とは,形質を目印に突然変異体を選抜する順遺伝学的選抜とは逆に,遺伝子を目印に突然変異体を選抜する方法である.例えば,変異原処理集団の全個体の全ゲノム配列を決定すれば,どの個体のどの遺伝子が機能欠損変異を有しているか予測することができる.全ゲノム配列の代わりに,TILLING 法(McCallum et al. 2000),High Resolution Melt(HRM) 解析 (Wittwer 2009) ,アンプリコンシーケンス (Tsuda et al. 2015) などの変異遺伝子検出技術を用いることで,特定の遺伝子に変異を持つ突然変異体を比較的容易に選抜することができる.このような逆遺伝学的選抜により VaMYB26 や rps1 責任遺伝子のダイズのオルソログの各パラログの突然変異体を選抜した後,交配により多重変異体を選抜すれば,裂莢が完全に抑制されたダイズを開発できる可能性がある.
雑豆の遺伝資源
ダイズの育種素材(breeding material)に利用できるのは Glycine 属だけであるが,ダイズの育種にあたって多様な雑豆に学ぶ価値は高く,ダイズの遺伝子源(gene source)ということになると,マメ科全ての遺伝資源(genetic resource)を含む.
近年,ダイズだけでなく雑豆でも,栽培化形質や環境適応形質に関する研究が世界的に盛んで,それらの遺伝資源が求められている.栽培化形質を研究する場合,対象となる種の栽培型と祖先野生種の遺伝資源が必要となる.例えば,上述したアズキの難裂莢性の研究では,徳島県で収集された在来品種(アクセッション番号:JP81481)とネパールで収集された祖先野生種(アクセッション番号:JP107881)が用いられた(Takahashi et al. 2020).一方,進化の過程で生じた環境適応形質を研究する場合には近縁野生種の遺伝資源が必要である.例えば,Vigna 属植物における乾燥耐性と塩害耐性の多様性を明らかにした研究では,ジーンバンクに保存されていた国内外の栽培型 9 種 15 アクセッションと近縁野生種 28 種 54 アクセッションが用いられた(Iseki et al. 2016, 2018).
農研機構のジーンバンクは,ダイズだけでなく,雑豆の遺伝資源も保存しており,一般に向けて遺伝資源を配布している(https://www.gene.affrc.go.jp/index_j.php).現在,配布可能なマメ科遺伝資源を約 2 万点保存しており,それらは約 8 千点の在来品種および約 5 千点の野生種を含む.品目としてダイズなど Glycine 属は全体の半数である約 1 万点を占め,残りはアズキなど Vigna 属約 8 千点,インゲンマメなど Phaseolus 属約千点,ラッカセイなど Arachis 属約 6 百点,エンドウマメなど Pisum 属約 2 百点,ソラマメなど Vicia 属約百点を含む.さらに未活用の雑豆として,キマメ(Cajanus cajan (L.) Millsp.),グラスピー(Lathyrus sativus L.),シカクマメ(Psophocarpus tetragonolobus (L.) DC.),ナタマメ(Canavalia gladiata (Jacq.) DC.),ハッショウマメ(Mucuna pruriens (L.) DC.),ヒヨコマメ(Cicer arietinum L.),フジマメ(Lablab purpureus (L.) Sweet),ホースグラム(Macrotyloma uniflorum (Lam.) Verdc.),ホワイトルーピン(Lupinus albus L.),レンズマメ(Lens culinaris Medik.)も保存されている.
おわりに
栽培化形質のうち種子の大型化に関しては,雑豆の遺伝子情報をダイズに応用したフィジビリティスタディがあるものの実用化には至っていない.ケツルアズキ(Vigna mungo (L.) Hepper)と呼ばれる雑豆の種子重が 69%増加した突然変異体の責任遺伝子として VmPPD 遺伝子が同定され,RNAi 法によりダイズのパラロガス遺伝子対(GmPPD1:Glyma.10g244400 と GmPPD2:Glyma.20g150000)の発現が抑制された形質転換体は,種子重が 53%増加した(Naito et al. 2017).しかし,ゲノム編集により作出された GmPPD のパラロガス遺伝子対の二重変異をもつ形質転換体は,最大でも種子重が 26%増加に留まったほか,シビアな表現型を示す系統では正常な種子が得られていない(Kanazashi et al. 2018).その種子重が 26%増加した系統は GmPPD1 がインフレーム変異,GmPPD2 がフレームシフト変異をもち,正常な種子が得られなかった系統は両遺伝子にフレームシフト変異をもつ.このように,パラロガス遺伝子対を機能欠損させれば目的の形質が発現するというわけでもなく,実用化には高いハードルがある.
しかも,ダイズは進化の過程で 2 度の全ゲノム重複を経験しているため,発現量の変化や組織特異性など発現制御が分化した 4 つ以上のパラログを保有することがある.このため,2 倍体の雑豆では 1 つの遺伝子を機能欠損させれば済むところ,ダイズでは 4 つのパラログを同時に機能欠損させる必要な場合もあろう.変異原処理集団から 4 つの遺伝子の機能欠損変異を逆遺伝学的に選抜して 4 重変異体を作成する過程は時間と労力を要するが,最近では,ゲノム編集により,2 つ以上のパラログを一挙に機能欠損させることも可能になった(Abe et al. 2019).
このように,困難は多くあるものの科学技術の向上につれて,雑豆に学ぶダイズの育種は加速すると考えられ,それを支える遺伝資源の継続的維持や拡充と共に,雑豆の基礎研究の重要性が増してくると著者は信じている.
利益相反
すべての著者は開示すべき利益相反はない.
謝辞
本研究は JSPS 科研費「21KK0272」の助成を受けた.模式図の作成に協力頂いた農研機構 遺伝資源研究センター 植物資源ユニットの有賀裕剛博士に感謝する.
引用文献
- 1) Abbo S, Pinhasi van-Oss R, Gopher A, Saranga Y, Ofner I and Peleg Z (2014) Plant domestication versus crop evolution: a conceptual framework for cereals and grain legumes. Trends Plant Science, 19(6): 351-360.
- 2) Abe F, Haque E, Hisano H, Tanaka T, Kamiya Y, Mikami M, Kawaura K, Endo M, Onishi K, Hayashi T and Sato K (2019) Genome-edited triple-recessive mutation alters seed dormancy in wheat. Cell Reports, 28(5): 1362-1369.e4.
- 3) Armon S, Efrati E, Kupferman R and Sharon E (2011) Geometry and mechanics in the opening of chiral seed pods. Science, 333(6050): 1726-1730.
- 4) Assefa T, Beebe SE, Rao IM, Cuasquer JB, Duque MC, Rivera M, Battisti A and Lucchin M (2013) Pod harvest index as a selection criterion to improve drought resistance in white pea bean. Field Crops Research, 148: 24-33.
- 5) Bhor TJ, Chimote VP and Deshmukh MP (2014) Inheritance of pod shattering in soybean [Glycine max (L.) Merrill]. Electronic Journal of Plant Breeding, 5: 671-676.
- 6) Caviness CE (1965) Effects of relative humidity on pod dehiscence in soybeans. Crop Science, 5: 511-513.
- 7) Di Vittori V , Bitocchi E, Rodriguez M , Alseekh S, Bellucci E, Nanni L, Gioia T, Marzario S, Logozzo G, Rossato M, De Quattro C, Murgia ML, Ferreira JJ, Campa A, Xu C, Fiorani F, Sampathkumar A, Fröhlich A, Attene G, Delledonne M, Usadel B , Fernie AR, Rau D and Papa R (2021) Pod indehiscence in common bean is associated with the fine regulation of PvMYB26. Journal of Experimental Botany, 72(5): 1617-1633.
- 8) Dong Y, Yang X, Liu J, Wang BH, Liu BL and Wang YZ (2014) Pod shattering resistance associated with domestication is mediated by a NAC gene in soybean. Nature Communications, 5, 3352.
- 9) Erb RM, Sander JS, Grisch R and Studart AR (2013) Self-shaping composites with programmable bioinspired microstructures. Nature Communications, 4, 1712.
- 10) Fuller DQ and Allaby R (2009) Seed dispersal and crop domestication: shattering, germination and seasonality in evolution under cultivation. In Østergaard L (ed), Annual plant reviews 38: Fruit development and seed dispersal, Blackwell Publishing Ltd., New Jersey, 238-295.
- 11) Fuller DQ, Denham T, Arroyo-Kalin M, Lucas L, Stevens CJ, Qin L, Allaby RG and Purugganan MD (2014) Convergent evolution and parallelism in plant domestication revealed by an expanding archaeological record. Proceedings of the National Academy of Sciences of the United States of America, 111(17): 6147-6152.
- 12) Funatsuki H, Ishimoto M, Tsuji H, Kawaguchi K, Hajika M and Fujino K (2006) Simple sequence repeat markers linked to a major QTL controlling pod shattering in soybean. Plant Breeding, 125: 195-197.
- 13) Funatsuki H, Suzuki M, Hirose A, Inaba H, Yamada T, Hajika M, Komatsu K, Katayama T, Sayama T, Ishimoto M and Fujino K (2014) Molecular basis of a shattering resistance boosting global dissemination of soybean. Proceedings of National Academy of Sciences of the United States of America, 111(50): 17797-17802.
- 14) Gladstones JS (1967) Selection for economic characters in Lupinus angustifolius and L. digitatus. Australian Journal of Experimental Agriculture and Animal Husbandry, 7: 360-366.
- 15) Iseki K, Takahashi Y, Muto C, Naito K and Tomooka N (2016) Diversity and evolution of salt tolerance in the genus Vigna. PLoS One, 11, e0164711.
- 16) Iseki K, Takahashi Y, Muto C, Naito K and Tomooka N (2018) Diversity of drought tolerance in genus Vigna. Frontiers in Plant Science, 9, 729.
- 17) Isemura T, Kaga A, Konishi S, Ando T, Tomooka N, Han OK and Vaughan DA (2007) Genome dissection of traits related to domestication in azuki bean (Vigna angularis) and comparison with other warm-season legumes. Annals of Botany, 100(5): 1053-1071.
- 18) Kaga A, Isemura T, Tomooka N and Vaughan DA (2008) The genetics of domestication of the azuki bean (Vigna angularis). Genetics, 178(2): 1013-1036.
- 19) Kaga A, Isemura T, Shu Y, Isobe S, Tabata S, Shimizu T, Somta P, Srinives P, Tomooka N and Vaughan D (2011) Asian Vigna genome research. Vth International Congress on Legume Genetics and Genomics: 106.
- 20) Kanazashi Y, Hirose A, Takahashi I, Mikami M, Endo M, Hirose S, Toki S, Kaga A, Naito K, Ishimoto M, Abe J and Yamada T (2018) Simultaneous site-directed mutagenesis of duplicated loci in soybean using a single guide RNA. Plant Cell Reports, 37(3): 553-563.
- 21) Kikuchi K, Koizumi M, Ishida N and Kano H (2006) Water uptake by dry beans observed by micro-magnetic resonance imaging. Annals of Botany, 98(3): 545-553.
- 22) Koizumi M, Kikuchi K, Isobe S, Ishida N, Naito S and Kano H (2008) Role of seed coat in imbibing soybean seeds observed by micro-magnetic resonance imaging. Annals of Botany, 102(3): 343-352.
- 23) Lo S, Muñoz-Amatriaín M, Boukar O, Herniter I, Cisse N, Guo YN, Roberts PA, Xu S, Fatokun C and Close TJ (2018) Identification of QTL controlling domestication-related traits in cowpea (Vigna unguiculata L. Walp). Scientific Reports, 8, 6261.
- 24) McCallum CM, Comai L, Greene EA and Henikoff S (2000) Targeting induced local lesions IN genomes (TILLING) for plant functional genomics. Plant Physiology, 123(2): 439-442.
- 25) Murgia ML, Attene G, Rodriguez M, Bitocchi E, Bellucci E, Fois D, Nanni L, Gioia T, Albani DM, Papa R and Rau D (2017) A comprehensive phenotypic investigation of the “Pod-Shattering Syndrome” in common bean. Frontiers in Plant Science, 8, 251.
- 26) Naito K, Takahashi Y, Chaitieng B, Hirano K, Kaga A, Takagi K, Ogiso-Tanaka E, Thavarasook C, Ishimoto M and Tomooka N (2017) Multiple organ gigantism caused by mutation in VmPPD gene in blackgram (Vigna mungo). Breeding Science, 67(2): 151-158.
- 27) Parker TA, Berny Mier Y Teran JC, Palkovic A, Jernstedt J and Gepts P (2020) Pod indehiscence is a domestication and aridity resilience trait in common bean. New Phytologist, 225: 558-570.
- 28) Parker TA, Lo S and Gepts P (2021) Pod shattering in grain legumes: emerging genetic and environment-related patterns. The Plant Cell, 33(2): 179-199.
- 29) Rau D, Murgia ML, Rodriguez M, Bitocchi E, Bellucci E, Fois D, Albani D, Nanni L, Gioia T, Santo D, Marcolungo L, Delledonne M, Attene G and Papa R (2019) Genomic dissection of pod shattering in common bean: mutations at non-orthologous loci at the basis of convergent phenotypic evolution under domestication of leguminous species. The Plant Journal, 97(4): 693-714.
- 30) Sakai H, Naito K, Ogiso-Tanaka E, Takahashi Y, Iseki K, Muto C, Satou K, Teruya K, Shiroma A, Shimoji M, Hirano T, Itoh T, Kaga A and Tomooka N (2015) The power of single molecule real-time sequencing technology in the de novo assembly of a eukaryotic genome. Scientific Reports, 5, 16780.
- 31) Schmutz J, Cannon SB, Schlueter J, Ma J, Mitros T, Nelson W, Hyten DL, Song Q, Thelen JJ, Cheng J, Xu D, Hellsten U, May GD, Yu Y, Sakurai T, Umezawa T, Bhattacharyya MK, Sandhu D, Valliyodan B, Lindquist E, Peto M, Grant D, Shu S, Goodstein D, Barry K, Futrell-Griggs M, Abernathy B, Du J, Tian Z, Zhu L, Gill N, Joshi T, Libault M, Sethuraman A, Zhang XC, Shinozaki K, Nguyen HT, Wing RA, Cregan P, Specht J, Grimwood J, Rokhsar D, Stacey G, Shoemaker RC and Jackson SA (2010) Genome sequence of the palaeopolyploid soybean. Nature, 463(7278): 178-183.
- 32) Sherwood S and Fu Q (2014) A drier future? Science, 343: 737-739.
- 33) Smýkal P, Vernoud V, Blair MW, Soukup A and Thompson RD (2014) The role of the testa during development and in establishment of dormancy of the legume seed. Frontiers in Plant Science, 5, 351.
- 34) 高橋 有,友岡憲彦 (2017) アズキがダイズを育種する. 国際農林業協力,40(1):28-35.
- 35) Takahashi Y, Sakai H, Yoshitsu Y, Muto C, Anai T, Pandiyan M, Senthil N, Tomooka N and Naito K (2019) Domesticating Vigna stipulacea: a potential legume crop with broad resistance to biotic stresses. Frontiers in Plant Science, 10, 1607.
- 36) Takahashi Y, Kongjaimun A, Muto C, Kobayashi Y, Kumagai M, Sakai H, Satou K, Teruya K, Shiroma A, Shimoji M, Hirano T, Isemura T, Saito H, Baba-Kasai A, Kaga A, Somta P, Tomooka N and Naito K (2020) Same locus for non-shattering seed pod in two independently domesticated legumes, Vigna angularis and Vigna unguiculata. Frontiers in Genetics, 11, 748.
- 37) Tsuda M, Kaga A, Anai T, Shimizu T, Sayama T, Takagi K, Machita K, Watanabe S, Nishimura M, Yamada N, Mori S, Sasaki H, Kanamori H, Katayose Y and Ishimoto M (2015) Construction of a high-density mutant library in soybean and development of a mutant retrieval method using amplicon sequencing. BMC Genomics, 16, 1014.
- 38) 上中登紀子,森 孝夫,豊沢 功 (2000) 豆類の吸水特性. 武庫川女子大学紀要,48:81-86.
- 39) Wang J, Sun P, Li Y, Liu Y, Yu J, Ma X, Sun S, Yang N, Xia R, Lei T, Liu X, Jiao B, Xing Y, Ge W, Wang L , Wang Z, Song X, Yuan M, Guo D, Zhang L, Zhang J , Jin D, Chen W, Pan Y, Liu T, Jin L, Sun J, Yu J, Cheng R, Duan X, Shen S, Qin J, Zhang MC , Paterson AH and Wang X (2017) Hierarchically aligning 10 legume genomes establishes a family-level genomics platform. Plant Physiology, 174(1): 284-300.
- 40) Watcharatpong P, Kaga A, Chen X and Somta P (2020) Narrowing down a major QTL region conferring pod fiber contents in yardlong bean (Vigna unguiculata), a vegetable cowpea. Genes, 11, 363.
- 41) Wittwer CT (2009) High-resolution DNA melting analysis: advancements and limitations. Human Mutation, 30(6): 857-859.
- 42) Yamada T, Makita H, Funatsuki H, Takahashi K, Hirata K, Hishinuma A and Tanaka J (2017) Causal analysis of yield-increase by introgression of shattering resistance gene pdh1 in soybean (in Japanese with English abstract). Japanese Journal of Crop Science, 86(3): 251-257.
- 43) Yoshimura Y, Mizuguti A and Matsuo K (2011) Analysis of the seed dispersal patterns of wild soybean as a reference for vegetation management around genetically modified soybean fields. Weed Biology and Management, 11: 210-216.
- 44) Zhang J and Singh AK (2020) Genetic control and geo-climate adaptation of pod dehiscence provide novel insights into soybean domestication. G3: Genes, Genomes, Genetics, 10: 545-554.