原著論文
長野県中部における断片化した小規模ブナ林の更新動態
2025 年 75 巻 2 号 p. 153-166

詳細
2025 年 75 巻 2 号 p. 153-166
長野県松本市牛伏寺において人為的に断片化された小規模ブナ林の更新動態を明らかにするため、林分構造とブナの種子生産特性を把握した。胸高断面積合計(調査面積0.555 ha、樹高≧3 m)は66.15 m2/haで、アカマツが48.2%、ブナが22.0%、コナラが18.0%を占めた。胸高直径分布はアカマツが25–85 cmに偏り、コナラは25–30 cmにピ一クをもつ一山型、ブナは95 cmまで連続的でL字型を示した。低木層(樹高0.5–3 m)ではブナ1個体のみが確認され、更新は停滞していた。これは、2000年頃までのスズタケによる被圧と、その後のシカ食害の影響と考えられる。稚樹(樹高<0.5 m)では、ナラ類(コナラ・ミズナラ)が全域に分布し、密度は0.47個体/m2であった。ブナ稚樹は全体の約3割の区画で確認されたが、密度は0.0176個体/m2と極めて低く、ほとんどが8年生以下であった。アカマツ稚樹はごく少数しか確認されなかった。2005–2024年の既報の種子モニタリングではブナの豊作はなく、並作が2回のみだった。先行研究が指摘する遺伝的多様性の低下により稔性が劣ると推察される。ブナの更新は困難であり、今後はナラ類が優占する林分への遷移が進むと予想されるが、シカの食害が続けばその更新も停滞する可能性がある。
We investigated the regeneration dynamics of a small, fragmented stand of Siebold’s beech (Fagus crenata) at Gofukuji Temple in Matsumoto, central Japan, in terms of stand structure and long-term seed production. The stand was 0.555 ha in area, and the total basal area of trees ≥3 m in height was 66.15 m2/ha. The stand was dominated by three species: Japanese red pine (Pinus densiflora; 48.2% of the total basal area); Siebold’s beech (22.0%); and Jolcham oak (Quercus serrata; 18.0%). Pines showed a broad range of diameter at breast height (DBH) values, at 25–85 cm, whereas oaks and beeches had unimodal and L-shaped DBH distributions that peaked at 25–30 and 95 cm, respectively. Only one beech individual was found in the understory (height: 0.5–3 m), which suggests poor regeneration, likely due to past suppression by dense dwarf bamboo (Sasa borealis) cover and ongoing sika deer (Cervus nippon) browsing. Among saplings (height:<0.5 m), oaks (Q. serrata and Quercus crispula) were widespread (density: 0.47 individuals/m2), whereas beech saplings were limited to 30% of the stand area (density: 0.0176 individuals/m2) and mainly aged ≤8 years, and red pine saplings were rare. An analysis of long-term seed monitoring data (2005–2024) detected no mast years and only two moderate seed crops. These results are consistent with previous genetic studies of this fragmented population in suggesting reduced seed viability due to inbreeding and pollen limitation. Given the pine population decline, likely caused by pine wilt disease, and the extremely poor beech regeneration in this stand, oak species may become dominant in the future, although continued deer browsing could eventually suppress forest regeneration overall.
ブナFagus crenata Blumeは、日本の温帯を代表する高木性の落葉広葉樹である。北海道南部から九州南部にかけて広く自生し、積雪量の多い日本海側では純林状の森林を形成する(Horikawa 1972; 藤田 1987)。しかし、第二次世界大戦時の木材需要や戦後の拡大造林による伐採、その後の土地利用開発などにより、ブナ林の分布域は大きく縮小・断片化し、現在は多くの林分が隔離分布している(環境庁 1988; 福嶋ほか 1995)。
人為的影響で断片化した林分では、母樹の減少や隔離により、特定母樹に依存した交配や自家受粉の確率が高まり、遺伝的多様性の低下などの影響が生じる可能性がある(Saunders et al. 1991; Lande 1998; Frankham et al. 2002)。一方、ブナのように長寿命で風媒の高木種は、花粉散布による遺伝子流動が大きく、異系交配の頻度も高いため、一般的な草本植物に比べて集団間の遺伝的分化は小さく、集団内の遺伝的多様性は高い傾向にある。そのため、断片化による影響を受けにくいと指摘されている(Hamrick and Godt 1996)。ところが、ブナの近縁種であるヨーロッパブナFagus sylvatica L.では、断片化した集団において遺伝的多様性の減少や近親交配の増加が生じており、それに伴って集団間の遺伝的分化が進行していることが報告されている(Jump and Peñuelas 2006)。このことから、断片化によって隔離された日本のブナ小集団においても、同様の影響が生じている可能性は否定できない。しかし、こうした林分における個体群の更新動態に関する知見は乏しいのが現状である。
本研究で対象としたブナ林(以下、牛伏寺ブナ林)は、長野県中部に位置する小規模な林分(集団)であり、過去の人為的な土地利用によって断片化され、特に他の集団から隔離されている(小山ほか 2002)。これまでに当ブナ林では、ブナ種子の発芽率が低い傾向にあること(小山・井田 2013)、および実生の多くが当林分内の成木由来であること(Inanaga et al. 2016)などが報告されており、近親交配や花粉制限に起因する遺伝的多様性の低下が示唆されている。また、小山ほか(2014)は、ブナの成木と下層個体群のサイズ構造に基づき、現時点では個体群が維持されているものの、下層個体の多くは約200年前から残る少数の母樹に由来する可能性を指摘している。
これらの先行研究は、主にブナ個体群の構造や遺伝的状態に焦点を当てたものであり、森林群落の持続性を考える上では不十分である。このため、ブナ以外の樹種や稚樹・実生の動態も含めた、群落全体としての更新過程の把握が必要である。特に、断片化によって将来的にブナの更新が制限される可能性がある場合、他樹種の更新や遷移の方向性を明らかにすることは、断片林分の長期的な種組成や構造の変化を理解し、森林群落全体の保全・管理方針を検討する上で重要である。例えば、同林分ではマツ枯れが進行しており、ブナ林分でも高い優占度を占めるアカマツPinus densiflora Siebold et Zucc.は、今後その割合が低下すると予測され、こうした他種の動態は森林の更新過程に大きな影響を及ぼすと考えられる。
さらに、ブナの種子生産量は年ごとに大きく変動することが知られており(Yasaka et al. 2003; Nielsen et al. 2004; Kon et al. 2005a; Masaki et al. 2008; Ida 2021)、この特性も林分の更新動態に大きく関与している。牛伏寺ブナ林では、著者の一人である井田が2005年以降、種子トラップを用いたブナ種子生産量の継続的なモニタリングを実施してきた。2024年までの20年間にわたるデータおよび調査手法の詳細は、Ida(2025)に報告されている。ただし、これまでのところ、種子生産特性に関する詳細な分析は行われていない。
以上を踏まえ、本研究では、小面積で断片化された牛伏寺ブナ林を対象に、その更新動態の解明を目的とする。特に、これまで詳細な検討が行われてこなかったブナ以外の構成樹種の個体群構造と、ブナの更新可能性を左右する主要な要因の一つである種子生産特性に着目する。具体的には、他樹種を含む稚樹から成木に至るまでのすべての発育段階を対象に個体群構造を把握し、林分の更新履歴を推定するとともに、2005年から2024年までの20年間にわたるブナ種子生産量モニタリングの既報データを用いて、その特性を明らかにする。本研究の成果は、断片化されたブナ林における更新の条件および制約要因に関する知見を提供し、類似する小規模ブナ林分の効果的な保全・管理の実践に資するものである。
牛伏寺ブナ林は、長野県松本市東部に位置する牛伏寺の寺有林の一部(36°09′56″N、138°01′10″E、標高990–1040 m)である。松本平の里地里山に残る唯一のブナ林として、学術的に貴重であり、長野県の希少植物群落(長野県 2005)および松本市特別天然記念物に指定されている。鉢伏山(標高 1929 m)の西側中腹に立地し、地質は第三紀の閃緑岩分布域にある(牛伏寺誌編纂委員会 2013)。調査区(後述)内の林冠下の2地点において、地表面から2 mの高さで実測した温度データ(井田 2025)によると、測定期間全体における年平均気温は10.2 ˚Cであった。月平均の日最高気温は8月の25.7°C、日最低気温は1月の−5.2 ˚Cであった。年間降水量は、北北西に約10 km離れた松本特別地域気象観測所(36°14′49″N、137°58′14″E、標高610 m)における平年値(1991–2020年)で1045.1 mmである。
冬期の最深積雪は、調査区内の1地点における実測値(井田 2025)によれば、2003/04年冬期から2020/21年冬期(18冬期)の平均は30 cmであった。また、根雪期間(積雪が3日以上継続して地表を覆う期間と定義)は、2005/06年冬期から2020/21年冬期までの16冬期のうち、最長で19日間(2013/14年冬期)であった。7冬期では根雪期間が3–8日間認められたが、残る8冬期では根雪は生じなかった。
牛伏寺ブナ林に近接するブナ集団(半径7 km以内)としては、東方約2 kmに2個体、約6 kmの地点に小規模な林分が確認されているのみである(小山ほか 2002; Inanaga et al. 2016)。当林分がいつ頃から残されてきたのかを示す資料は見つかっておらず、また植栽の記録も存在しない。ただし、戦国時代の1534年に牛伏寺が現在の地に移転した記録はある(牛伏寺誌編纂委員会 2013)。その後、江戸時代末期(1860年頃)には濫伐や山火事により一帯の山林が荒廃し、それを受けて明治18(1885)年から大正7(1918)年にかけて、周囲の河川で大規模な砂防工事が行われた(牛伏川砂防工事沿革史編纂会 1933)。
小山ほか(2014)によると、牛伏寺ブナ林における現在のブナの林冠木(樹高20 m以上)の樹齢は少なくとも200年程度と推定されており、これに基づけば1860年頃の時点で、既に40年生程度のブナ林分が寺院周辺に孤立して成立していたと考えられている。なお、Koyama et al.(2012)による遺伝的構造の解析からは、当ブナ集団が日本海側系統と太平洋側系統の中間に位置づけられることが示されている。
現在、ブナ個体群は牛伏寺の寺院周辺のみに分布している。小山ほか(2014)によれば、寺院の北側に位置する丘陵状の尾根部から南向きの急斜面(平均傾斜30°)にかけて、東西約110 m、南北約60 mの範囲に、樹高1.2 m以上のブナが100個体(83株)確認されている。また、それとは別に、寺院の南側に位置する標高1000–1010 mの尾根(寺院から約120–130 m南方)でも、6個体(4株)のブナが確認されている。本研究では、当地域のブナ集団の存続を大きく左右すると考えられる前者のブナ林分の更新過程について検討を行った。なお、2021年現在、調査対象としたブナ林分の周囲は、コナラ二次林、スギCryptomeria japonica(L.f.)D.DonやヒノキChamaecyparis obtusa(Siebold et Zucc.)Endl.の植林地、およびモウソウチクPhyllostachys edulis(Carrière)Houz.からなる竹林に囲まれている。ブナ林分とこれら縁辺林分との間では、林冠高が概ね連続しており、林分境界における下層植生に大きな違いは見られない。
調査区は、牛伏寺の寺院南側に分布する6個体(4株)を除いた、寺院北側に成立するブナ林分の全個体群を対象として、2021年4–5月に設置した。稚樹(実生を含む)から成木に至るすべての発育段階のブナ個体が含まれるよう、水平距離で5 mまたは10 mごとに境界杭(十全株式会社製エタプロンK-35赤)を設置しながらコンパス測量を実施した。その結果、10 m×10 m区画が56個、10 m×5 m区画が3個となり、総面積は0.575 haとなった。
この林分には、アカマツとコナラQuercus serrata Murrayの高木が混交しているが、アカマツは2019年頃からマツ枯れにより一部が伐採処理されている。著者(井田)の観察によれば、林床は2000年頃までは桿高2 m近くに達するスズタケSasa borealis(Hack.)Makino et Shibataが全域に繁茂していたが、ニホンジカCervus nippon Temminck(以下、シカ)による摂食圧により2010年頃までにその植被のほとんどが失われ、現在は部分的に残るのみとなっている(図1)。今回の調査時(2021年6–10月)に記録したスズタケの植被率は、5 m×5 m区画230個の平均値(±標準偏差)で2.8±5.6%、平均高は11.8±6.3 cmであった。林床の草本類はわずかに出現するのみであり、特に高木種の定着や生育への影響はほとんどないと考えられることから、本研究では調査対象外とした。

樹木個体群の構造を把握するため、樹高3 m以上の上層木を対象に毎木調査を実施した。スギ植林地区画(0.02 ha)を対象外としたため、調査区の総面積は0.555 haである。今後も恒久的に樹木の追跡調査を行うため、各樹幹には地上1.35 mの位置に、個体識別用の番号を刻印したアルミタグをステンレス釘で取り付けた。
毎木調査では、樹種名と階層(高木層:樹高10 m以上、亜高木層:樹高3 m以上10 m未満)を記録し、高さ1.3 mにおける周囲長(胸高周囲長)をスチールメジャーで0.1 cm単位で測定した。根元で複数の萌芽幹が出ている場合や、胸高より下部で分枝している場合には、それぞれが同株(同一個体)であることを記載し、個々の樹幹を調査対象とした。これらの調査は2021年8月および10月に実施した。
データの集計にあたっては、胸高周囲長を円周率で除して胸高直径を算出し、胸高断面積合計(調査区での総計と、単位面積あたりの合計値[1 ha換算])を求めた。
樹高3 m未満の低木層(単幹は樹高0.5 m以上、萌芽幹は0.5 m未満も含む)については、幹ごとに樹種名と樹高を記録した。樹高の測定には、3 mの測量ポールを用いた。
高木種稚樹の分布調査今後の森林動態を推定するため、主要な高木種の稚樹(実生も含む)の生育状況を調べた。ここでは、上記の毎木調査の対象外である樹高0.5 m未満の個体(萌芽幹を除く)を「稚樹」と定義し、当年–数年生の実生を含め、樹種ごとの分布と個体密度を把握した。
調査は、10 m×10 mおよび10 m×5 mの区画をそれぞれ5 m×5 m区画に4および2分割し、スギ植林地の区画(0.02 ha[8区画])も含めた合計230区画において実施した。スギ植林区を含めたのは、その一部でブナ稚樹が確認されたためである。
稚樹の調査対象種は、上層木の胸高断面積合計で優占していた上位3種(アカマツ、ブナ、コナラ[ここでは後述のとおり「ナラ類」として扱った])に、230区画のうち36区画(調査区の東端域、標高990–1100 m)で実施した予備調査(2021年4–5月、調査区設置時に実施)において、2区画以上で稚樹の定着が確認された高木種3種(ウラジロモミAbies homolepis Siebold et Zucc.、クリCastanea crenata Siebold et Zucc.、ケヤキZelkova serrata(Thunb.)Makino)を加えた計6樹種とした。
予備調査では、本調査に向けた調査デザインの検討および稚樹の種組成の傾向を把握することを目的とし、調査効率の観点から、個体数を樹種別に以下の6段階で記録した:0(なし)、1(1–5個体)、2(6–15個体)、3(16–30個体)、4(31–60個体)、5(61個体以上)。
調査区内には、ミズナラQuercus crispula Blume var. crispulaの成木が4個体確認されており(表1参照)、稚樹にもミズナラと判断される個体がいくらか見られた。しかし、コナラとの形態的識別が困難な場合があったため、集計および後述の解析においては、両種を区別せず「ナラ類」として扱った。
| 種名 | 幹数(本/0.555 ha) | 幹密度 | 胸高断面積合計 | ||||
|---|---|---|---|---|---|---|---|
| 高木層 | 亜高木層 | 合計 | 本/ha | % | m2/ha | % | |
| アカマツ | 56 | — | 56 | 100.9 | 9.6 | 31.88 | 48.19 |
| ブナ | 48 | 49 | 97 | 174.8 | 16.6 | 14.52 | 21.95 |
| コナラ | 60 | 14 | 74 | 133.3 | 12.7 | 11.89 | 17.98 |
| ヤマモミジ | 8 | 20 | 28 | 50.5 | 4.8 | 1.94 | 2.93 |
| ヒノキ(植栽) | 5 | 5 | 10 | 18 | 1.7 | 1.31 | 1.98 |
| サワシバ | 1 | 39 | 40 | 72.1 | 6.9 | 0.71 | 1.07 |
| ケアオダモ | 2 | 109 | 111 | 200 | 19 | 0.6 | 0.9 |
| アオハダ | 2 | 64 | 66 | 114.8 | 11.3 | 0.59 | 0.89 |
| ミズナラ | 3 | 1 | 4 | 7.2 | 0.7 | 0.51 | 0.77 |
| イタヤカエデ | 2 | — | 2 | 3.6 | 0.3 | 0.51 | 0.77 |
| ウラジロモミ | 2 | 5 | 7 | 12.6 | 1.2 | 0.38 | 0.57 |
| スギ(植栽) | 6 | 6 | 12 | 21.6 | 2.1 | 0.33 | 0.5 |
| ミズキ | 2 | — | 2 | 3.6 | 0.3 | 0.25 | 0.38 |
| アワブキ | 1 | 37 | 38 | 68.5 | 6.5 | 0.17 | 0.26 |
| シナノキ | 2 | 3 | 5 | 9 | 0.9 | 0.16 | 0.24 |
| ケヤキ | 1 | — | 1 | 1.8 | 0.2 | 0.09 | 0.14 |
| ウリカエデ | — | 14 | 14 | 25.2 | 2.4 | 0.08 | 0.13 |
| コブシ | — | 1 | 1 | 1.8 | 0.2 | 0.05 | 0.08 |
| クリ | 1 | — | 1 | 1.8 | 0.2 | 0.04 | 0.07 |
| ヤマボウシ | 1 | — | 1 | 1.8 | 0.2 | 0.04 | 0.06 |
| オオヤマザクラ | — | 2 | 2 | 3.6 | 0.3 | 0.03 | 0.05 |
| アズキナシ | — | 5 | 5 | 9 | 0.9 | 0.03 | 0.05 |
| ソヨゴ | — | 5 | 5 | 9 | 0.9 | 0.03 | 0.04 |
| ウラゲエンコウカエデ | — | 1 | 1 | 1.8 | 0.2 | <0.01 | 0.01 |
| ナツハゼ | — | 1 | 1 | 1.8 | 0.2 | <0.01 | <0.01 |
| 総計 | 203 | 381 | 584 | 1048.1 | 100 | 66.15 | 100 |
同年6月から10月にかけて実施した本調査では、残りの194区画において、主要高木種の更新状況をより定量的に評価するため、稚樹の個体数を樹種ごとにカウントして記録した。
なお、ブナ稚樹については、予備調査区を含むすべての調査区画で個体識別(油性ペンで番号を記したプラスチックプレートを針金で枝に固定)を行い、齢構成の推定を目的として、樹齢の指標とされる芽鱗痕数(Nakashizuka 1983)を記録した。
ブナ種子生産量の評価本研究では、牛伏寺ブナ林において2005年から著者の一人(井田)により実施されてきた、種子トラップを用いた種子落下量のモニタリングデータ(Ida 2025)を用いて、当林分におけるブナの種子生産特性を把握した。特に、更新に寄与するのは発芽可能な健全種子であることから、年ごとの健全種子数(個/m2)および健全種子率(種子総数[雌花数]に対する健全種子数の比率)に着目して評価を行った。また、種子の豊凶を評価するため、各年の健全種子数を中島・小谷(2013)の基準に従って、豊作(下限値200個/m2)、並作(同50個/m2)、不作(同5個/m2)、凶作(5個/m2未満)の4区分に分類した。
統計解析主要高木種(ブナ、ナラ類[コナラおよびミズナラ]、アカマツ)における上層木(樹高3 m以上)と稚樹(樹高0.5 m未満)との関係は、一般化線形混合モデル(GLMM)により解析した。GLMMでは、応答変数を各樹種の5 m×5 m区画あたりの稚樹個体数、説明変数を同区画あたりの各樹種の胸高断面積合計値とし、誤差構造にはポアソン分布、リンク関数にはlogを、ランダム効果には区画ID(全230区画)を用いた。
各樹種について、すべての説明変数の組み合わせ(nullモデルも含む)に基づいてモデルを構築し、赤池情報量規準(AIC)を求めた。AICが最小のモデルとの差(ΔAIC)が2未満であるモデルを有効モデルとし、各樹種における胸高断面積と稚樹個体数の関係を検討した。
ここで、予備調査区(36区画)では、主要高木種のうちナラ類稚樹のみ個体数を階級で記録していた(ブナ稚樹は個体識別、アカマツ稚樹は出現なし)ため、GLMMにおいてはこれらの区画の応答変数として代替値を使用した。代替値は、ナラ類稚樹の個体数をカウントした他の194区画のデータを各階級(0–5)に変換し、階級ごとの平均個体数を算出して得たものである。ただし、階級5に該当する予備調査区は存在しなかった。階級1–4に対応する平均個体数は次の通りである。1:3個体(該当区画数9)、2:10個体(17)、3:22個体(3)、4:39個体(1)。階級0に該当する区画は6区画あった。
なお、代替値の使用が解析結果に及ぼす影響を検討するため、予備調査区(36区画)を除外した194区画のみを用いたGLMM解析もあわせて実施した。
統計解析はR 4.4.1(R Core Team 2024)を使用し、GLMMの構築にはlme4パッケージ、モデル選択にはMuMInパッケージを用いた。
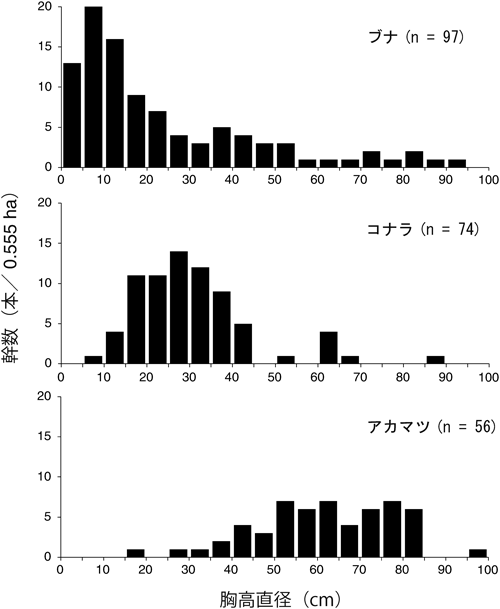
毎木調査から、樹高3 m以上の立木は25種、胸高断面積合計は66.15 m2/haであった(表1)。胸高断面積合計の中で各樹種の占める割合は、高い順にアカマツ(48.2%)、ブナ(22.0%)、コナラ(18.0%)であり、その他の樹種はいずれも5%未満であった。幹数は、これら主要高木種では多い順にブナ(174.8本/ha、16.6%)、コナラ(133.3本/ha、12.7%)、アカマツ(100.9本/ha、9.6%)となっていた。アカマツはすべて樹高10 m以上の高木であり、亜高木(樹高3 m以上10 m未満)は存在しなかった。これら主要高木種の胸高直径サイズ分布(図2)を見ると、ブナは95 cmまで幅広く連続的で10 cm未満の多いL字型を示した。コナラは5–45 cmに集中しており、25–30 cmにピ一クをもつ一山型を示した。アカマツは25–85 cmに連続的に分布し中・大径木に偏っていた。空間的には、ブナが存在しない場所にコナラやアカマツが分布し、アカマツは尾根筋で優占していた(図3)。

低木層(樹高0.5 m以上3 m未満)では、調査面積0.555 haにおいて計12樹種229本が確認された(表2)。なかでも、アオハダIlex macropoda Miq.(69本)、ケアオダモFraxinus lanuginose Koidz. f. lanuginosa(58本)、アワブキMeliosma myriantha Siebold et Zucc.(44本)の出現本数が多く、これらは亜高木層(表1)でも多く見られた。しかし、これらの樹種が全体に占める胸高断面積合計の割合は1%未満と小さかった(表1)。一方、高木層で優占していたアカマツやコナラは低木層では確認されず、ブナは1個体(樹高1.8 m、0.00018個体/m2)のみであった。
| 種名 | 樹高≧2 m | 樹高≧0.5 m、 <2 m | 萌芽幹 (樹高<0.5 m) | 総計 |
|---|---|---|---|---|
| アオハダ | 31 | 37 | 1 | 69 |
| ケケアオダモ | 30 | 26 | 2 | 58 |
| アワブキ | 19 | 23 | 2 | 44 |
| サワシバ | 6 | 9 | 3 | 18 |
| ナツハゼ | 7 | 6 | — | 13 |
| ダンコウバイ | 1 | 5 | 2 | 8 |
| ヤマツツジ | 1 | 5 | — | 6 |
| ウラジロモミ | — | 5 | — | 5 |
| ヤマモミジ | 2 | 1 | — | 3 |
| ウリカエデ | 2 | — | — | 2 |
| フジ | — | 2 | — | 2 |
| ブナ | — | 1 | — | 1 |
| 総計 | 99 | 120 | 10 | 229 |
稚樹(樹高0.5 m未満、調査面積0.575 ha)では、ナラ類(コナラおよびミズナラ)が最も多く、213区画(全体の92.6%)に出現し、比較的高密度で分布していた(表3、図3)。次いで、ウラジロモミが81区画(35.2%)、ブナが66区画(28.7%)、クリが24区画(10.4%)で確認された。アカマツとケヤキの稚樹も確認されたが、出現区画は全体の5%未満であった。ブナ稚樹の区画あたりの出現数はすべて5個体未満であり、全体では101個体(0.0176個体/m2)が確認された。
| 種名 | 階級*(個体数:本/25 m2) | 出現区画数(n=230) | |||||
|---|---|---|---|---|---|---|---|
| 1 (1–5) | 2 (6–15) | 3 (16–30) | 4 (31–60) | 5 (61–) | 個 | % | |
| ナラ類 (コナラ・ミズナラ) | 64 | 98 | 35 | 13 | 3 | 213 | 92.6 |
| ウラジロモミ | 80 | 1 | — | — | — | 81 | 35.2 |
| ブナ | 66 | — | — | — | — | 66 | 28.7 |
| クリ | 24 | — | — | — | — | 24 | 10.4 |
| アカマツ | 10 | 1 | — | — | — | 11 | 4.8 |
| ケヤキ | 9 | 1 | — | — | — | 10 | 4.3 |
5 m×5 m区画を単位として、ブナ、ナラ類、アカマツの上層木の胸高断面積と各樹種の稚樹密度との関係をGLMMにより解析した結果、ブナ稚樹の密度については、3樹種いずれの上層木の胸高断面積とも明瞭な関係は認められなかった(表4)。ΔAICが2未満の有効モデルにはアカマツやナラ類を説明変数に含むものもあったが、いずれも切片のみが有意であった。
| 応答変数 | モデル順位 | (切片) | 係数 | AIC | ΔAIC | ||
|---|---|---|---|---|---|---|---|
| ブナ | ナラ類 | アカマツ | |||||
| ブナ稚樹 | 1 | −1.426 | — | — | — | 255.77 | 0.00 |
| 2 | −1.407 | — | — | −0.254 | 257.66 | 1.90 | |
| 3 | −1.436 | — | 0.354 | — | 257.72 | 1.96 | |
| 4 | −1.432 | 0.139 | — | — | 257.75 | 1.99 | |
| ナラ類稚樹 | 1 | 1.929 | 2.510 | — | 763.41 | 0.00 | |
| 2 | 1.964 | −0.878 | 2.429 | — | 763.79 | 0.38 | |
| 3 | 1.898 | 2.586 | 0.364 | 764.61 | 1.19 | ||
| 4 | 2.102 | −0.880 | 2.504 | 0.363 | 764.99 | 1.57 | |
一方、ナラ類稚樹の密度に対しては、すべての有効モデルにおいてナラ類の上層木が正の効果を示した。一部のモデルではブナやアカマツも説明変数に含まれていたが、これらの係数は有意ではなかった。また、アカマツ稚樹を応答変数とするモデルでは、出現個体数が計24個体(11区画)と極めて少なかったため、モデルは収束しなかった。
なお、予備調査区(36区画)を除いた194区画のみを用いて実施したGLMM解析においても、ナラ類稚樹に対するナラ上層木の正の効果は一貫して認められた。これにより、稚樹個体数における代替値の使用が解析結果に及ぼす影響は限定的であることが確認された。具体的には、第3・第4モデルのAIC順位が入れ替わったのみであり、最良モデルおよびその解釈に影響はなかった。
調査区内で確認されたブナ稚樹101個体の芽鱗痕数は、2–8個の範囲に分布し、最大は12個であった(図4)。このことから、大半は8年生以下、すなわち概ね2012年以降に定着したと推定される。なかでも37個体と最も多かった芽鱗痕数3個の個体群は、並作年であった2017年の種子に由来する可能性が高い。

ブナの種子生産量は、モニタリング期間(2005–2024年)において、年ごとの総数が最大474個/m2(2005年)から最小0.8個/m2(2014年)までの範囲で変動した(Ida 2025)。健全種子数に基づく豊作年は、20年間の調査期間中に一度も確認されず、並作年は2回(2017、2022年)にとどまった(図5)。不作年は6回(2006、2011、2015、2018、2020、2024年:5.6–24.0個/m2)、凶作年は12回を数えた。

健全種子率が最も高かったのは2017年の29.5%であり、この年は健全種子数も最大(128.4個/m2)であった。次いで健全種子率が高かった2022年(25.4%)においても、健全種子数は92.2個/m2と2番目に多かった。
牛伏寺ブナ林では、胸高断面積合計においてアカマツ、ブナ、コナラの順に優占し、これら3樹種で全体の約9割を占めていた(表1)。樹高3 m以上のブナは、胸高直径分布の範囲が広く、L字型のサイズ構造を示したことから(図2)、長期にわたって継続的または断続的な更新が繰り返されてきたこと、すなわち定常的に新規個体が加入し続けたことが示唆される。一方、一山型のサイズ分布を示したコナラは、かつての薪炭利用の放棄に伴い、実生や萌芽により短期間に一斉更新が生じた可能性がある。また、アカマツの胸高直径分布は中・大径木が主体であったことから、コナラの薪炭利用放棄時よりもさらに以前に定着した個体が長期的に成長してきたと考えられる。これらのことは、里山としての林分利用履歴と自然更新の相互作用により現在の林分構造が形成されたことを示しており、小山ほか(2014)の見解を支持する結果である。
ブナについては同様の手法による調査で既報と一致した結果が得られたが、本研究では、アカマツやコナラも、現在の林分構造および今後の更新動態に大きな影響を及ぼす主要な構成樹種であることが新たに示された。
低木層発達の停滞要因低木層(樹高0.5以上3 m未満)ではアカマツやナラ類は確認されず、ブナもわずか1個体のみであった(表2)。このように低木層がほとんど発達していないことは、稚樹の成長や個体の更新が一時的に停滞していた可能性を示している。
本林分では、2000年前後を境に、林床を広く覆っていたスズタケがシカの摂食圧により衰退し始めたことが確認されている(図1)。一般に、スズタケのようなササ類は、高木種の実生や稚樹の定着・成長を妨げることが知られており(Nakashizuka 1988; Ida and Nakagoshi 1996; Abe et al. 2001)、スズタケが繁茂していた時期(2000年頃以前)には、実生の更新や稚樹バンクの形成が制限されていたと考えられる。このため、現在、低木層より上層に達している個体群は、当時スズタケの被圧を既に抜けていたか、その葉群下でわずかに生き残っていた個体に由来すると推察される。
一方、当林分におけるコナラの一斉更新は、牛伏寺ブナ林周辺がかつて里山として利用されていた背景を踏まえると、全国的に薪炭利用の放棄が進んだ1970年前後に生じた可能性がある。スズタケも、こうした里山管理の下では採草や踏圧などの影響を受けて、当時の優占度はそれほど高くなかったと推察される。これにより、比較的開放的な林床環境の下で、コナラに加えてブナも更新し、現在の低木層より上層を構成する個体群が成立したと考えられる。
2000年頃以降は、スズタケの衰退によって林床の被度が低下した一方で、並行してシカによる直接的な摂食圧が稚樹にも及ぶようになったと推察される。田村(2013)は、シカ食害によりスズタケが退行した太平洋側のブナ林において、植生保護柵の外ではブナを含む高木種の稚樹の樹高が0.1 m以下に抑えられていたと報告している。また、米山ほか(2010)は、ササ刈り地におけるブナ植栽木がニホンカモシカCapricornis crispus Temminckにより樹高0.6–0.8 mで集中的に採食され、1 m以上では頂端部の採食が見られなかったことを示しており、シカにおいても同様の傾向が想定される。
一般に、ササ類は数十年から百数十年に一度、一斉に開花して枯死する特異な生活史を持ち、一斉枯死後には林床が明るくなることで高木種の稚樹の定着が一時的に促進される(例えば、Nakashizuka 1988; Abe et al. 2001)。しかし本林分では、2000年以降のスズタケの衰退は一斉枯死ではなくシカによる摂食が主因であった。そのため、林床が開放的になったにもかかわらず、同時に稚樹も食害を受けるようになり、結果として低木層の発達は停滞したと考えられる。
以上を踏まえると、現在の低木層が未発達な状況は、スズタケによる被圧と、それに続くシカによる摂食圧によって、稚樹から低木層への移行が一時的に妨げられてきたことを反映しているといえる。
稚樹の更新状況稚樹では、ナラ類(コナラ・ミズナラ)が最も高密度かつ広範囲に分布していた(表3、図3)。GLMMの解析結果(表4)では、ナラ類の稚樹密度は、同種の上層木の胸高断面積に正の影響を受けており、その樹冠下においてより多くの稚樹が定着していることが示唆された。すなわち、ナラ類においては、一斉更新により成立した上層木(母樹)の樹幹下で、種子(堅果)の散布から稚樹の定着に至る更新プロセスが、林分全体で機能していることが示唆される。なお、ミズナラは上層木が4個体存在し(表1)、稚樹も確認されたが、同種と識別できた稚樹の出現数は多くはなかったことから、更新への寄与はコナラが中心となると考えられる。
ブナの稚樹は一定数確認されたものの、その出現頻度はナラ類に比べて低く、密度は0.0176個体/m2であった。これは、太平洋側ブナ林の東京都三頭山における0.0345個体/m2(島野・沖津 1993)を大きく下回り、日本海側のブナ林において報告されている、ササのない林分での1.2個体/m2(3–8年生)、林冠ギャップ下での1.6個体/m2(2–12年生)(Nakashizuka and Numata 1982a, b)と比べても、極めて低い値であった。
また、芽鱗痕数の分布(図4)からは、調査区内に現存するブナ稚樹の多くが、過去10年程度の種子生産に由来していることが示唆され、更新が近年に偏っていることがうかがえる。なお、芽鱗痕の読み取りには誤差が伴い、生育期の二次伸長により実際よりも多く記録される場合もある(小山 2016)。
GLMM解析では、ブナ稚樹密度とブナ・ナラ類・アカマツいずれの上層木との明瞭な関連性は認められなかった。この結果は、ブナの稚樹定着が特定の上層木の分布に依存せず、林内の比較的広い範囲で偶発的に生じていることを示唆する。これは、ブナ個体群の樹冠が林分の広域を覆っており、種子散布が林内全体に及んでいることに起因する。
一方、実生の定着および稚樹の生残や成長には、林冠構造よりも林床の微気候に左右されると推察される。現在の林分は低木層やササ類、草本層が発達しておらず、林床は比較的開放的である。こうした環境下では日射、温度、および乾燥の影響を受けやすく、種子サイズがナラ類より小さいブナは、乾燥や凍結による死亡リスクがより高い。特に根雪期間が短い当地のような寡雪環境では、積雪による保温・保湿効果が乏しく、種子や稚樹の越冬生残率は多雪地より大きく低下する(Shimano and Masuzawa 1998; Homma et al. 1999; 石井ほか 2007)。
加えて、2000年頃から顕著となったシカの摂食圧(図1)は、稚樹や低木層の個体への直接的な影響に加え、林床のスズタケの衰退を通じて林床環境全体を大きく変化させた。一方で、こうした変化により、スズタケによる被圧で妨げられていた稚樹の定着が一時的に促進された可能性もある(田村 2013; Itô et al. 2014)。実際、ナラ類の稚樹の多さはその影響と考えられ、2008年以降にはブナ稚樹の出現数がわずかに増加する傾向も確認された(図3)。しかし、こうした間接的な促進効果は長期的には持続せず、種子の乾燥や凍結、さらにシカによる摂食など、複合的な制約が継続しているため、定着後の生残は依然として厳しいと考えられる。
その他の樹種では、ウラジロモミの稚樹出現頻度がブナを上回っており、高木層に5個体、亜高木層に2個体(表1)、低木層に5個体(表2)が確認されたことから、小規模ながらも更新が断続的に進行していることがうかがえる。アカマツは、高木・亜高木層で優占しているものの、稚樹の出現頻度は極めて低く、当林分における今後の更新の見込みは低い。クリやケヤキについても稚樹が確認されたが、局所的であり、高木・亜高木層での個体数もそれぞれ1個体と少ないため、今後の群落構成への影響は限定的である。
ブナ種子の生産特性と稔性低下の要因牛伏寺ブナ林におけるブナ種子の生産特性は、豊作が数年ごとに訪れるという一般的なブナ林の傾向とは大きく異なり、20年間の調査期間中(Ida 2025)で豊作年は一度も確認されなかった。並作年も2回にとどまり、不作年は6回、凶作年は12回に及んだ。このことは、長期にわたり健全種子の供給が十分に行われていないことを示している。また、年間の種子総数(雌花数)が多かった2005、2011、2013年はいずれも不作年に分類され、並作年であった2017年と2022年も、他地域の林分と比べて健全種子率はそれぞれ29.5、25.4%と決して高いとはいえなかった(寺澤ほか 1995; 井田ほか 2017; Ida et al. 2023)。このような傾向は、健全種子率の変動が雌花数だけでなく、他の生理・生態学的要因の影響を受けていると考えられる。
ブナの健全種子形成に影響を与える要因としては、これまでに気温(Kon et al. 2005b; Kon and Noda 2007)や昆虫による種子捕食圧(Igarashi and Kamata 1997; Kon et al. 2005a)が指摘されてきた。しかし、牛伏寺ブナ林のように断片化された小規模な林分では、これらに加えて遺伝的要因の影響が生じている可能性がある。
ブナは自家不和合性を備えているものの、それは不完全であり(寺沢・柳井 1991; 寺澤ほか 1995)、隔離された小集団では他家受粉の機会が限られることで自殖や近親交配が増加し、遺伝的多様性の低下が懸念される(Ohkawa et al. 1998; Kitamura et al. 2008, 2015)。Inanaga et al.(2016)は、牛伏寺ブナ集団とその周辺2集団を対象に行った遺伝解析により、実生(2011年産の種子由来)の94%が牛伏寺集団内の成木を両親としていたことを示し、花粉制限が生じている可能性とともに、牛伏寺集団の健全種子率が長野県内の他の集団よりも低く、胚が未発達な種子の比率が高いことを報告している。また、同集団の健全種子における発芽率も相対的に低い傾向があった(小山・井田 2013)。牛伏寺ブナ林は、本州中央部に分布する小規模集団(成木が100個体未満の林分)の中でも、断片化後の世代経過期間が比較的長く、集団内の遺伝的多様性が低いことが示唆されている(福田・戸丸 未発表)。
以上のことから、牛伏寺ブナ林では、花粉供給の制限や遺伝的要因による稔性低下が、健全種子の減少をもたらしていると考えられる。
牛伏寺ブナ林の今後の推移現時点では、低木層より上層に生育する比較的若齢のブナ個体群が将来的に成長し林冠を形成することで、当面はブナ林の相観が維持されると考えられる。しかし、これまでの調査結果から、ブナの更新個体群は特定母樹に由来している可能性が高く、交配相手が限られていることから、集団内の遺伝的多様性のさらなる低下が懸念される。それに伴い、健全種子の生産量が今後さらに減少し、長期的にはブナ個体群の維持が困難になる可能性がある。
アカマツについては、尾根筋やギャップなどを除いては更新がほとんど生じておらず、マツ枯れの進行とともに林分での優占度は次第に低下すると予想される。
一方、コナラ中径木の優占と、林内に広く分布するナラ類(コナラおよびミズナラ)の高い稚樹密度は、今後、ナラ類がアカマツやブナに替わって林分を優占する可能性を示している。しかしながら、とりわけシカの摂食圧が継続する場合、ブナの稚樹定着や生残が引き続き阻害されるだけでなく、ナラ類を含めた低木層の発達も抑制され、森林全体の更新動態が停滞する可能性も否定できない。
本研究を実施するにあたり、金峰山牛伏寺の皆様には多大なる協力をいただきました。現地調査では信州大学教育学部森林生態学研究室の皆様の協力をいただきました。種子生産量のモニタリングでは原田佳一氏、宮﨑政幸氏、桜井智子氏、松田貴子氏の協力を得ました。調査の許認可については松本市教育委員会、信州大学研究推進部研究支援課および同大学教育学部森林生態学研究室の宮川文美氏にお世話になりました。以上の方々にこの場を借りて感謝申し上げます。本研究の一部はJSPS科研費(25292087、23H02256)の助成を受けたものである。