Abstract
Tumor necrosis factor α (TNF-α), a pro-inflammatory cytokine, regulates inflammatory and immune responses by up-regulating gene expression in a manner that is dependent on the transcription factor nuclear factor κB (NF-κB). In the present study, we found that 4-hydroxypanduratin A and isopanduratin A, constituents of the rhizomes of Boesenbergia pandurata, inhibited the TNF-α-stimulated up-regulation of intercellular adhesion molecule-1 (ICAM-1) in human lung adenocarcinoma A549 cells. 4-Hydroxypanduratin A and isopanduratin A also reduced ICAM-1 mRNA expression and NF-κB-responsive luciferase activity in TNF-α-stimulated A549 cells. Moreover, 4-hydroxypanduratin A and isopanduratin A prevented the TNF-α-stimulated translocation of the NF-κB subunit p65 to the nucleus and the phosphorylation and proteasomal degradation of the inhibitor of the NF-κB α protein. The present results revealed that 4-hydroxypanduratin A and isopanduratin A inhibit TNF-α-stimulated gene expression and the NF-κB-dependent signaling pathway in A549 cells.
INTRODUCTION
Pro-inflammatory cytokines activate multiple transcription factors, including nuclear factor κB (NF-κB), which up-regulates many target genes that are essential for inflammatory and immune responses.1) In response to pro-inflammatory cytokines, such as tumor necrosis factor α (TNF-α), cell adhesion molecules, including intercellular adhesion molecule-1 (ICAM-1) and vascular cell adhesion molecule-1 (VCAM-1), are up-regulated on vascular endothelial cells, and regulate the adhesion and transmigration of circulating lymphocytes into inflammation sites.2,3) Upon a TNF-α stimulation, TNF receptor 1 induces the activation of inhibitor of NF-κB (IκB) kinases, which phosphorylate IκB associating with NF-κB dimers in the cytoplasm.4) Phosphorylated IκB subsequently undergoes ubiquitination and proteasome degradation.5) After dissociating from IκB, the NF-κB heterodimer, composed of p65 (also known as RelA) and p50, enters the nucleus, and activates the transcription of NF-κB target genes.1,6)
Panduratins are cyclohexene chalcones belonging to the flavonoid family.7) Panduratin A and its derivatives have been shown to exert various biological effects, including anti-tumor, anti-viral, and anti-inflammatory activities.7–9) We previously reported the isolation of eight new cyclohexene chalcones and seven known related compounds, e.g., panduratin A, 4-hydroxypanduratin A (Fig. 1A), and isopanduratin A (Fig. 1A), from the methanol extract of the rhizomes of Boesenbergia pandurata.10) Consistent with previous findings,11) panduratin A, 4-hydroxypanduratin A, and isopanduratin A have been shown to exhibit anti-tumor activity against human pancreatic cancer PANC-1 cells.10) Previous studies demonstrated that panduratin A inhibited the NF-κB signaling pathway in the murine monocyte cell line RAW264.7 and human lung adenocarcinoma A549 cells.12–14) The expression of NF-κB was reduced by panduratin A in lipopolysaccharide (LPS)-induced human gingival fibroblast-1 cells.15) However, in contrast to panduratin A, the biological activities of 4-hydroxypanduratin A and isopanduratin A on the NF-κB signaling pathway remain unclear. In the course of our screening, we found that 4-hydroxypanduratin A and isopanduratin A prevented the TNF-α-stimulated up-regulation of ICAM-1 in A549 cells. In the present study, we elucidated the biological activities of 4-hydroxypanduratin A and isopanduratin A on TNF-α-stimulated gene expression and the NF-κB-dependent signaling pathway.
MATERIALS AND METHODS
CellsHuman lung adenocarcinoma A549 cells (JCRB0076) were provided by the National Institutes of Biomedical Innovation, Health and Nutrition JCRB Cell Bank (Osaka, Japan). A549 cells were maintained as previously described.16,17)
Reagents4-Hydroxypanduratin A and isopanduratin A were purified as the (±)-form from the methanol extract of the rhizomes of Boesenbergia pandurata, as described in our previous study10) and used in this study. The extraction and isolation of cardamonin were described in the Supplementary materials. Recombinant human TNF-α was provided by Dainippon Pharmaceuticals (Osaka, Japan).
Cell Enzyme-Linked Immunosorbent Assay (ELISA)The amount of ICAM-1 on the cell surface was assessed by cell ELISA, as described in our previous studies.16,17)
Cell Viability AssayCells were treated with 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT) for the last 2 h of the incubation. The MTT assay was performed as described in our previous studies.16,17)
Quantitative PCRThe amounts of ICAM-1 mRNA and β-actin mRNA were measured by real-time PCR. The preparation of RNA, cDNA synthesis, and real-time PCR were described previously.18)
Luciferase Reporter AssayThe luciferase reporter assay using the NF-κB-responsive luciferase reporter plasmid (a kind gift from Dr. Ralph C. Budd) was performed as described previously.18,19)
Confocal Microscopic ObservationsA549 cells were grown on coverslips coated with Cellmatrix® Type I-C (Nitta Gelatin, Osaka, Japan). A549 cells were fixed with 4% paraformaldehyde–phosphate-buffered saline (PBS) and permeabilized with 0.3% Triton X-100–PBS, followed by an incubation overnight with 5% normal goat serum–PBS. Cells were reacted serially with a mouse anti-p65 antibody (2A12A7; Thermo Fisher Scientific, Waltham, MA, U.S.A.) as the primary antibody and Alexa Fluor 488-conjugated anti-mouse immunoglobulin G (IgG) antibody (Thermo Fisher Scientific) as the second antibody, followed by staining with Hoechst 33342. The confocal laser scanning microscope system FV10i (Olympus, Tokyo, Japan) was used to analyze stained cells.
Western BlottingCell lysate preparation and Western blotting were described previously.16,17) An anti-IκBα antibody (25; BD Biosciences, San Jose, CA, U.S.A.), anti-phospho-IκBα (Ser32/36) antibody (5A5; Cell Signaling Technology, Danvers, MA, U.S.A.), anti-VCAM-1 antibody (E1E8X, Cell Signaling Technology), and anti-β-actin antibody (2F3; Wako Pure Chemical Industries, Ltd., Osaka, Japan) were used as primary antibodies. ImageQuant LAS 4000 mini (GE Healthcare Japan, Tokyo, Japan) was used to analyze and quantify protein bands.
Statistical AnalysisA one-way ANOVA and Tukey’s test was used in statistical analyses for multiple comparisons.
RESULTS AND DISCUSSION
A549 cells were treated with 4-hydroxypanduratin A and isopanduratin A for 1 h and then stimulated with TNF-α for 6 h. The amount of cell-surface ICAM-1 was markedly up-regulated in response to the TNF-α stimulation. TNF-α-stimulated ICAM-1 expression was inhibited by 4-hydroxypanduratin A and isopanduratin A at 25–50 µM (Figs. S1A, B). Under the same incubation conditions, the cell viability of A549 cells was assessed by the MTT assay. 4-Hydroxypanduratin A at concentrations up to 50 µM did not affect cell viability (Fig. S1C), while isopanduratin A decreased cell viability at 50 µM (Fig. S1D).
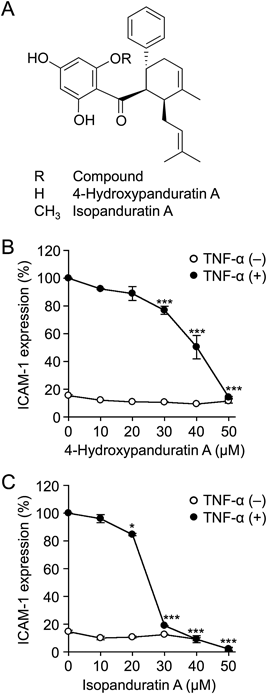
To further clarify whether 4-hydroxypanduratin A and isopanduratin A inhibit TNF-α-stimulated ICAM-1 expression in a dose-dependent manner, A549 cells were treated with doses of 10-µM increments for 1 h and then stimulated with TNF-α for 6 h. 4-Hydroxypanduratin A and isopanduratin A inhibited TNF-α-stimulated ICAM-1 expression in a dose-dependent manner and almost completely at concentrations of 50 and 30–50 µM, respectively (Figs. 1B, C). We also investigated whether the pretreatment with 4-hydroxypanduratin A and isopanduratin A was needed for the inhibition of TNF-α-stimulated ICAM-1 expression. The results obtained showed that the pretreatment was not needed for the inhibition of TNF-α-stimulated ICAM-1 expression by 4-hydroxypaduratin A and isopanduratin A (Figs. S2A, B).
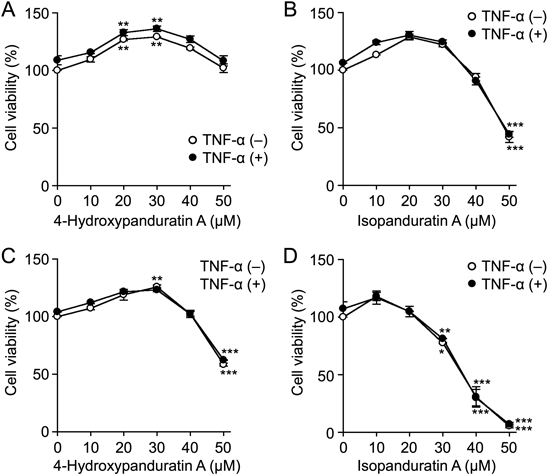
The MTT assay showed that 4-hydroxypanduratin A and isopanduratin A did not markedly decrease cell viability at concentrations up to 50 and 40 µM, respectively, during a 7-h incubation (Figs. 2A, B). These results, together with Figs. 1B and C, indicate that 4-hydroxypaduratin A and isopanduratin A inhibited the TNF-α-stimulated up-regulation of ICAM-1, at least at concentrations of 50 and 30–40 µM, respectively, without decreasing cell viability. In contrast, 4-hydroxypanduratin A and isopanduratin A decreased cell viability at concentrations of 50 and 40–50 µM, respectively, during a 25-h incubation (Figs. 2C, D). These results showed that 4-hydroxypaduratin A and isopanduratin A exhibited increasing cytotoxic activities with longer treatment durations.
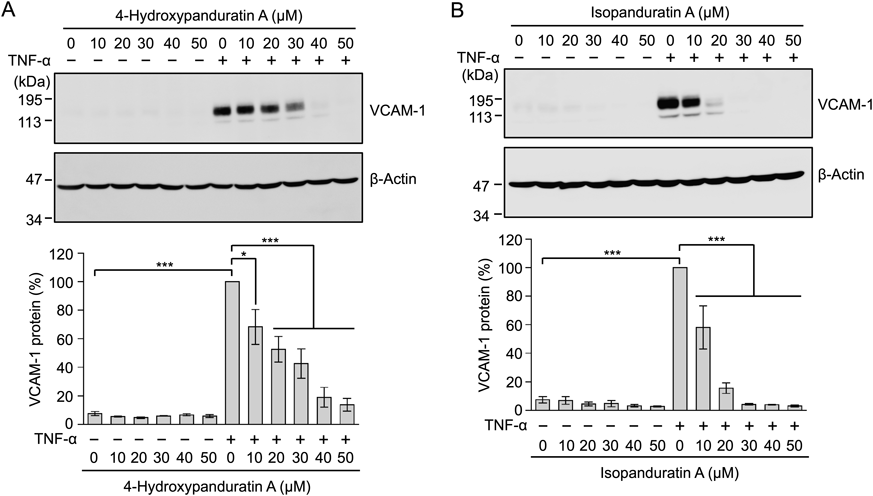
In addition to ICAM-1, VCAM-1 is up-regulated by proinflammatory cytokines, such as TNF-α.20) We previously reported that the expression of VCAM-1 was up-regulated by an interleukin-1α stimulation in A549 cells.21) A549 cells were treated with 4-hydroxypanduratin A and isopanduratin A for 1 h and then stimulated with TNF-α for 6 h. Cell lysates were prepared and analyzed by Western blotting. 4-Hydroxypanduratin A and isopanduratin A inhibited the TNF-α-stimulated up-regulation of the VCAM-1 protein at lower concentrations than those that inhibited ICAM-1 expression (Fig. 3).
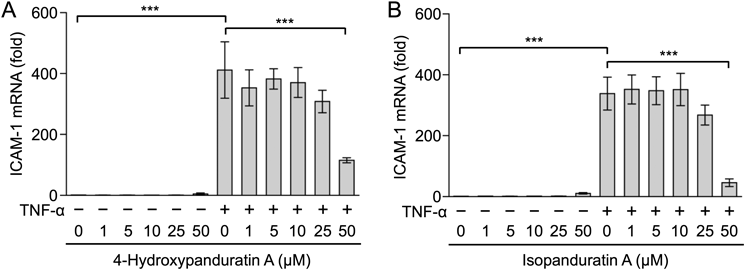
We then investigated the effects of 4-hydroxypanduratin A and isopanduratin A on ICAM-1 mRNA expression. A549 cells were treated with 4-hydroxypanduratin A and isopanduratin A for 1 h and then stimulated with TNF-α for 2 h. The TNF-α stimulation markedly up-regulated the expression of ICAM-1 mRNA (Figs. 4A, B). 4-Hydroxypanduratin A and isopanduratin A inhibited the TNF-α-stimulated up-regulation of ICAM-1 mRNA at a concentration of 50 µM (Figs. 4A, B).
The ICAM-1 gene is mainly activated by the NF-κB transcription factor in response to TNF-α.22,23) NF-κB-responsive luciferase reporters were used to evaluate effects on NF-κB-dependent transcription activity. The 2.5-h stimulation with TNF-α enhanced luciferase activity by approximately 4-fold (Figs. 5A, B). 4-Hydroxypanduratin A at 50 µM and isopanduratin A at 25–50 µM inhibited TNF-α-stimulated NF-κB-responsive luciferase activity (Figs. 5A, B). Under these conditions, 4-hydroxypanduratin A was not cytotoxic, while isopanduratin A exhibited slight cytotoxicity against A549 cells (Figs. 5C, D). These results suggested that 4-hydroxypanduratin A and isopanduratin A inhibited NF-κB-dependent gene expression.
Cardamonin (Fig. S3A) is a chalcone derivative that is structurally related to panduratins. It has been reported to inhibit the NF-κB signaling pathway.24,25) Hence, cardamonin was used as a known inhibitor targeting the NF-κB signaling pathway. We showed that cardamonin inhibited TNF-α-induced ICAM-1 expression in a dose-dependent manner (Fig. S3B), but did not affect cell viability under these conditions (Fig. S3C).
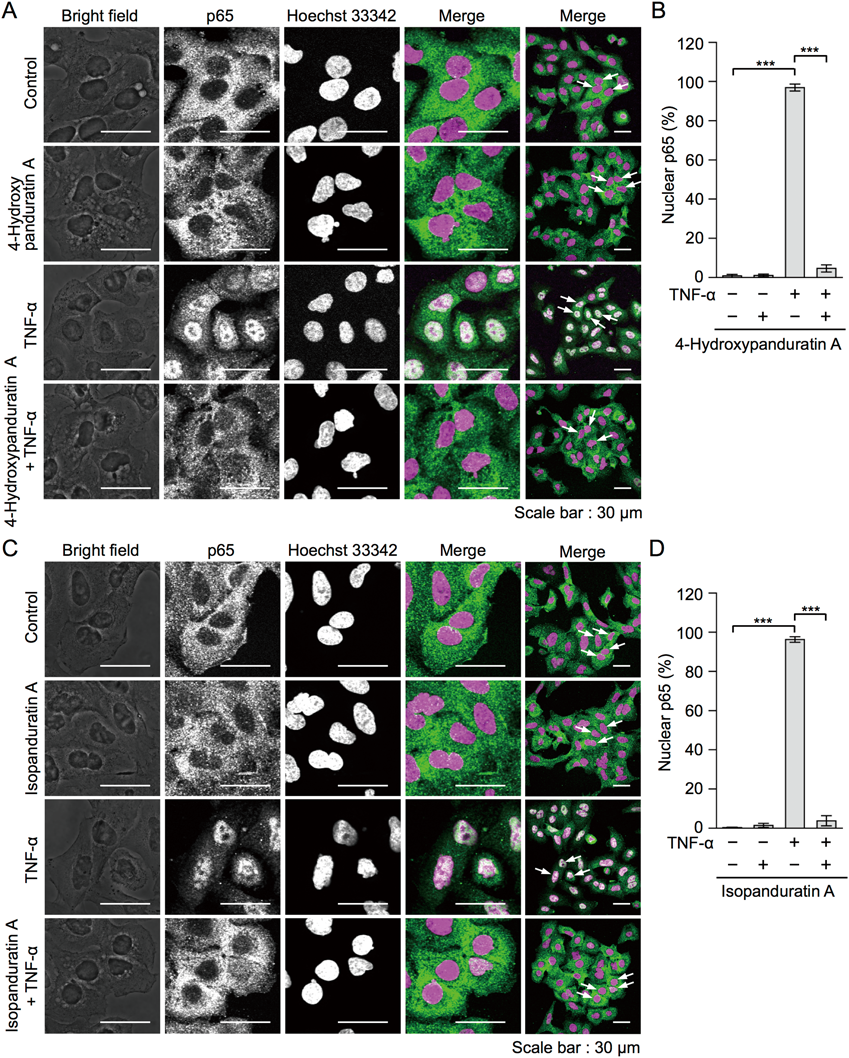
The NF-κB subunit p65 plays a major role in the TNF-α-dependent NF-κB signaling pathway.4,5) The p65 and p50 heterodimers are present in the cytoplasm by interacting with IκBα. A549 cells were treated with 4-hydroxypanduratin A and isopanduratin A for 1 h and then stimulated with TNF-α for 30 min, followed by immunostaining for p65 and nuclear staining. p65 mainly localized to the cytoplasm in unstimulated A549 cells, whereas the translocation of p65 to the nucleus was induced by the TNF-α stimulation (Figs. 6A–D). 4-Hydroxypanduratin A at 50 µM and isopanduratin A at 25 µM inhibited the TNF-α-stimulated nuclear translocation of p65 (Figs. 6A–D).
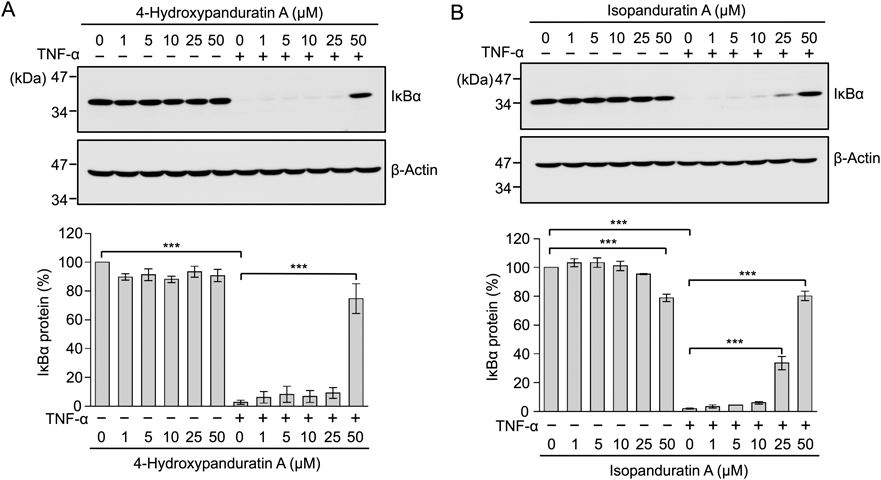
The IκB kinase complex phosphorylates IκBα, and phospho-IκBα is immediately poly-ubiquitinated and subsequently hydrolyzed by the proteasome, prior to the translocation of p65 to the nucleus.4,5) A549 cells were treated with 4-hydroxypanduratin A and isopanduratin A for 1 h and then stimulated with TNF-α for 15 min. The amount of IκBα was almost completely diminished 15 min after the TNF-α stimulation (Figs. 7A, B). 4-Hydroxypanduratin A at 50 µM and isopanduratin A at 25–50 µM inhibited the TNF-α-stimulated degradation of IκBα (Figs. 7A, B).
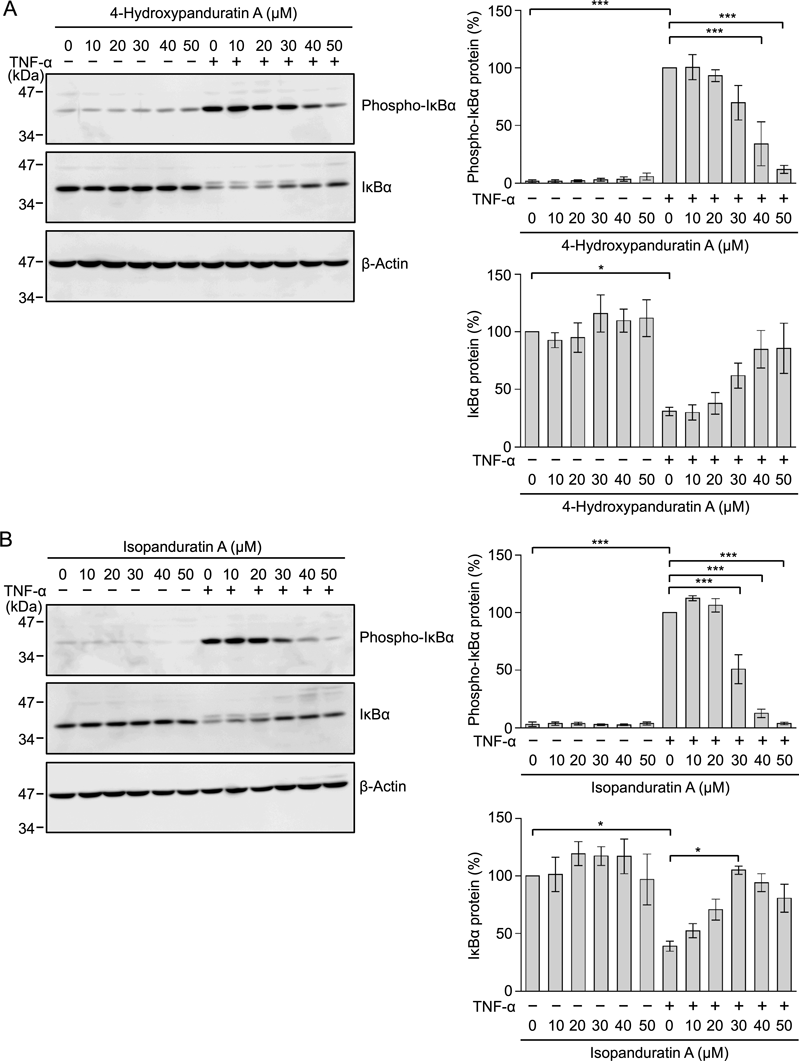
To examine their effects on TNF-α-induced IκBα phosphorylation, A549 cells were treated with 4-hydroxypanduratin A and isopanduratin A for 1 h and then stimulated with TNF-α for 5 min. 4-Hydroxypanduratin A and isopanduratin A inhibited TNF-α-stimulated IκBα phosphorylation in a dose-dependent manner (Figs. 8A, B). These results indicated that 4-hydroxypanduratin A and isopanduratin A inhibited the IκB kinase complex or its upstream steps in the NF-κB signaling pathway.
Panduratin A has been shown to inhibit LPS-induced IκBα phosphorylation and its degradation in RAW264.7 cells,12) as well as the TNF-α-stimulated translocation of p65 and p50 to the nucleus in A549 cells.13,14) A previous study reported that panduratin A reduced the expression of NF-κB in LPS-stimulated gingival fibroblast-1 cells.15) Collectively, these findings indicated that panduratin A inhibited the NF-κB signaling pathway. However, to the best of our knowledge, previous studies did not provide direct evidence to show that 4-hydroxypanduratin A and isopanduratin A inhibited the NF-κB signaling pathway, except that this may be related to the NO inhibitory activities of 4-hydroxypanduratin A.26) The present results clearly indicate that 4-hydroxypanduratin A and isopanduratin A inhibited the NF-κB signaling pathway upon the TNF-α stimulation.
4-Hydroxypanduratin A and isopanduratin A exerted inhibitory effects on the NF-κB signaling pathway at 50 and 25–50 µM, respectively. 4-Hydroxypanduratin A at 50 µM did not affect cell viability for up to 7 h, but decreased it by 25 h, whereas isopanduratin A did not markedly affect cell viability at 50 µM up to 2.5 h and reduced it at 40–50 µM by 25 h. These results showed that the cytotoxic activities of 4-hydroxypanduratin A and isopanduratin A increased slightly with longer incubation times. It appears that 4-hydroxypanduratin A and isopanduratin A block cellular targets at particular concentrations without affecting cellular viability during upregulation of the NF-κB and ICAM-1 signaling pathways. However, the range of effectiveness of 4-hydroxypanduratin A and isopanduratin A on the NF-κB signaling pathway and its downstream gene expression may be narrower than that of their cytotoxic activities. A structure–activity relationship study is needed to obtain more preferable panduratin derivatives as anti-inflammatory agents targeting the NF-κB signaling pathway.
The TNF-α-stimulated VCAM-1 expression was inhibited by 4-hydroxypanduratin A and isopanduratin A at lower concentrations, compared with those that inhibited ICAM-1 expression. The NF-κB transcription factor plays a major role in the up-regulation of ICAM-1.22,23) It has been reported that the expression of ICAM-1 and VCAM-1 is differently regulated in the differentiation of Caco-2 cells.27) Thus, panduratin derivatives may affect additional cellular processes other than the NF-κB signaling pathway. Further experiments are necessary to clarify this issue.
Panduratin A and its derivatives have been shown to exhibit anti-tumor activity toward a number of cancer cell lines.10,11,13,14,28,29) In contrast, panduratin A, 4-hydroxypanduratin A, and isopanduratin A inhibited TNF-α plus actinomycin D-induced cell death in L929 cells.30) Panduratin A and 4-hydroxypanduratin A have also been shown to reduce L-glutamate toxicity in N18-RE-105 cells.31) Thus, in addition to its anti-tumor activities, panduratin derivatives are assumed to possess biological activities capable of preventing cell death, although these may be cell context-specific effects. The further elucidation of the cellular targets of panduratin derivatives is needed to clarify the molecular mechanisms underlying their diverse biological activities.
Acknowledgments
This work was supported partly by the Japan Society for the Promotion of Science (JSPS) KAKENHI Grant number 16H04910 (to T.K.).
Conflict of Interest
The authors declare no conflict of interest.
Supplementary Materials
The online version of this article contains supplementary materials.
REFERENCES
- 1) Karin M, Greten FR. NF-κB: linking inflammation and immunity to cancer development and progression. Nat. Rev. Immunol., 5, 749–759 (2005).
- 2) Gerhardt T, Ley K. Monocyte trafficking across the vessel wall. Cardiovasc. Res., 107, 321–330 (2015).
- 3) Vestweber D. How leukocytes cross the vascular endothelium. Nat. Rev. Immunol., 15, 692–704 (2015).
- 4) Hayden MS, Ghosh S. Regulation of NF-κB by TNF family cytokines. Semin. Immunol., 26, 253–266 (2014).
- 5) Sasaki K, Iwai K. Roles of linear ubiquitinylation, a crucial regulator of NF-κB and cell death, in the immune system. Immunol. Rev., 266, 175–189 (2015).
- 6) Baud V, Karin M. Is NF-κB a good target for cancer therapy? Hopes and pitfalls. Nat. Rev. Drug Discov., 8, 33–40 (2009).
- 7) Rozmer Z, Perjési P. Naturally occurring chalcones and their biological activities. Phytochem. Rev., 15, 87–120 (2016).
- 8) Wahab HA, Yusof R, Rahman NA. A search for vaccines and therapeutic for Dengue: A review. Curr. Comput. Aided Drug Des., 3, 101–112 (2007).
- 9) Zhang EH, Wang RF, Guo SZ, Liu B. An update on antitumor activity of naturally occurring chalcones. Evid. Based Complement. Alternat. Med., 2013, 815621 (2013).
- 10) Nguyen NT, Nguyen MTT, Nguyen HX, Dang PH, Dibwe DF, Esumi H, Awale S. Constituents of the rhizomes of Boesenbergia pandurata and their antiausterity activities against the PANC-1 human pancreatic cancer line. J. Nat. Prod., 80, 141–148 (2017).
- 11) Win NN, Awale S, Esumi H, Tezuka Y, Kadota S. Bioactive secondary metabolites from Boesenbergia pandurata of Myanmar and their preferential cytotoxicity against human pancreatic cancer PANC-1 cell line in nutrient-deprived medium. J. Nat. Prod., 70, 1582–1587 (2007).
- 12) Yun JM, Kwon H, Hwang JK. In vitro anti-inflammatory activity of panduratin A isolated from Kaempferia pandurata in RAW264.7 cells. Planta Med., 69, 1102–1108 (2003).
- 13) Cheah SC, Appleton DR, Lee ST, Lam ML, Hadi AHA, Mustafa MR. Panduratin A inhibits the growth of A549 cells through induction of apoptosis and inhibition of NF-kappaB translocation. Molecules, 16, 2583–2598 (2011).
- 14) Cheah SC, Lai SL, Lee ST, Hadi AHA, Mustafa MR. Panduratin A, a possible inhibitor in metastasized A549 cells through inhibition of NF-kappa B translocation and chemoinvasion. Molecules, 18, 8764–8778 (2013).
- 15) Kim H, Kim MB, Kim C, Hwang JK. Inhibitory effects of panduratin A on periodontitis-induced inflammation and osteoclastogenesis through inhibition of MAPK pathways in vitro. J. Microbiol. Biotechnol., 28, 190–198 (2018).
- 16) Yamada Y, Taketani S, Osada H, Kataoka T. Cytotrienin A, a translation inhibitor that induces ectodomain shedding of TNF receptor 1 via activation of ERK and p38 MAP kinase. Eur. J. Pharmacol., 667, 113–119 (2011).
- 17) Mitsuda S, Yokomichi T, Yokoigawa J, Kataoka T. Ursolic acid, a natural pentacyclic triterpenoid, inhibits intracellular trafficking of proteins and induces accumulation of intercellular adhesion molecule-1 linked to high-mannose-type glycans in the endoplasmic reticulum. FEBS Open Bio., 4, 229–239 (2014).
- 18) Harada M, Morimoto K, Kondo T, Hiramatsu R, Okina Y, Muko R, Matsuda I, Kataoka T. Quinacrine inhibits ICAM-1 transcription by blocking DNA binding of the NF-κB subunit p65 and sensitizes human lung adenocarcinoma A549 cells to TNF-α and the Fas ligand. Int. J. Mol. Sci., 18, 2603 (2017).
- 19) Matsuda I, Matsuo K, Matsushita Y, Haruna Y, Niwa M, Kataoka T. The C-terminal domain of the long form of cellular FLICE-inhibitory protein (c-FLIPL) inhibits the interaction of the caspase 8 prodomain with the receptor-interacting protein 1 (RIP1) death domain and regulates caspase 8-dependent nuclear factor κB (NF-κB) activation. J. Biol. Chem., 289, 3876–3887 (2014).
- 20) Carter RA, Wicks IP. Vascular cell adhesion molecule 1 (CD106): A multifaceted regulator of joint inflammation. Arthritis Rheum., 44, 985–994 (2001).
- 21) Hiramatsu R, Fukuhara S, Mitsuda S, Yokomichi T, Kataoka T. Betulinic acid and oleanolic acid, natural pentacyclic triterpenoids, interfere with N-linked glycan modifications to intercellular adhesion molecule-1, but not its intracellular transport to the cell surface. Eur. J. Pharmacol., 767, 126–134 (2015).
- 22) Collins T, Read MA, Neish AS, Whitley MZ, Thanos D, Maniatis T. Transcriptional regulation of endothelial cell adhesion molecules: NF-κB and cytokine-inducible enhancers. FASEB J., 9, 899–909 (1995).
- 23) Roebuck KA, Finnegan A. Regulation of intercellular adhesion molecule-1 (CD54) gene expression. J. Leukoc. Biol., 66, 876–888 (1999).
- 24) Lee JH, Jung HS, Giang PM, Jin X, Lee S, Son PT, Lee D, Hong YS, Lee K, Lee JJ. Blockade of nuclear factor-κB signaling pathway and anti-inflammatory activity of cardamonin, a chalcone analog from Alpinia conchigera. J. Pharmacol. Exp. Ther., 316, 271–278 (2006).
- 25) Israf DA, Khaizurin TA, Syahida A, Lajis NH, Khozirah S. Cardamonin inhibits COX and iNOS expression via inhibition of p65NF-κB nuclear translocation and Iκ-B phosphorylation in RAW264.7 macrophage cells. Mol. Immunol., 44, 673–679 (2007).
- 26) Tewtrakul S, Subhadhirasakul S, Karalai C, Ponglimanont C, Cheenpracha S. Anti-inflammatory effects of compounds from Kaempferia parviflora and Boesenbergia pandurata. Food Chem., 115, 534–538 (2009).
- 27) Astarci E, Sade A, Çimen I, Savaş B, Banerjee S. The NF-κB target genes ICAM-1 and VCAM-1 are differentially regulated during spontaneous differentiation of Caco-2 cells. FEBS J., 279, 2966–2986 (2012).
- 28) Yun JM, Kwon H, Mukhtar H, Hwang JK. Induction of apoptosis by panduratin A isolated from Kaempferia pandurata in human colon cancer HT-29 cells. Planta Med., 71, 501–507 (2005).
- 29) Yun JM, Kweon MH, Kwon H, Hwang JK, Mukhtar H. Induction of apoptosis and cell cycle arrest by a chalcone panduratin A isolated from Kaempferia pandurata in androgen-independent human prostate cancer cells PC3 and DU145. Carcinogenesis, 27, 1454–1464 (2006).
- 30) Morikawa T, Funakoshi K, Ninomiya K, Yasuda D, Miyagawa K, Matsuda H, Yoshikawa M. Medicinal foodstuffs. XXXIV. Structures of new prenylchalcones and prenylflavanones with TNF-α and aminopeptidase N inhibitory activities from Boesenbergia rotunda. Chem. Pharm. Bull., 56, 956–962 (2008).
- 31) Shindo K, Kato M, Kinoshita A, Kobayashi A, Koike Y. Analysis of antioxidant activities contained in the Boesenbergia pandurata SCHULT. Rhizome. Biosci. Biotechnol. Biochem., 70, 2281–2284 (2006).