日本では1980年代以降ニホンジカCervus nippon(以下、シカ)の個体数増加と分布拡大に伴い、農林業や自然植生に深刻な被害が生じている。シカ個体群動態は対象となる地域の気候、食物利用可能性、捕獲圧等様々な要因によって変動しうることが知られているため、対象地域に生息する個体群の齢構成や妊娠率等の基礎的な情報の把握は、地域の実態に即した適切な管理計画を策定する上で重要である。本研究では、2000年代に分布が拡大し個体数が増加したと考えられる長野県浅間山周辺地域で捕獲されたシカ301頭を対象に、齢構成、繁殖および成長特性に関する基礎情報を明らかにすることを目的とした。対象集団の年齢構成を調査した結果、メスの年齢構成に占める若齢個体(0–1歳)の割合は経年的な増加傾向を示したことから、生息密度の増加に伴う幼獣生存率の低下が生じている可能性は低く、集団が依然として高い繁殖力を維持していることが示唆された。繁殖特性を評価するために妊娠率を算出した結果、本調査地域の性成熟齢(1歳)における妊娠率(50.8%)は他地域と比較するとやや低いと考えられた一方で、成獣(2歳以上)の妊娠率は91.0%であった。成長特性を評価するために体重および体サイズ指標の一つである頭蓋骨最大長の成長曲線を算出した結果、体重はメスで約5歳まで、オスで約12歳まで増加し、頭蓋骨最大長はメスで約3–4歳、オスで約7–8歳頃まで増加すると予測された。他地域と比較すると、本調査地域の集団は個体の成長が遅れており、最大値に到達するまでの時間が長いと考えられた。本調査地域では上述の通り、幼獣生存率の低下が生じている可能性が低く、成獣が高い妊娠率を維持していることから、個体群の増加を抑制できていない可能性が高い。一方で近年の高密度化に伴い、初産齢の遅延や体重および体サイズの成長速度の低下が生じている可能性がある。個体数の更なる増加を抑制するためには引き続き高い捕獲圧をかける必要があるとともに、捕獲個体の分析によるモニタリングを継続して行い、その変化を評価する必要がある。
Since the 1980s, population and distribution expansions of sika deer (Cervus nippon) in Japan have caused serious damage to agriculture, forestry and the natural environment. Deer population dynamics are affected by various factors including climate, food availability and hunting pressure, which vary regionally. Therefore, fundamental information describing local populations, such as age structure and fecundity, is important for developing appropriate management plans. We assessed the age structure, reproduction and body growth of 301 deer killed in the Mt. Asama region of Nagano Prefecture, Japan, where deer populations and distributions have expanded since the 2000s. The proportion of young females (aged 0–1 year) tended to increase over time, suggesting that juvenile survival may not decrease as population density increases. The study population appeared to maintain high pregnancy rates, with that of yearlings (50.8%) slightly lower than those of other local populations, whereas adult females (aged>2 years) had a higher pregnancy rate (91.0%). Growth curves based on body weight attained a plateau in females and males at 5 and 12 years of age, respectively, whereas those based on the total skull length, a body size indicator, plateaued in females and males at 3–4 and 7–8 years, respectively. Therefore, compared to other local populations, the deer population in the Mt. Asama region is expected to grow more slowly and take longer to reach maximum body weight and size. Because juvenile survival is not decreasing and adult females are maintaining high pregnancy rates, this deer population may still be expanding. However, recent population density increases may have delayed the age at first pregnancy and slowed growth in terms of body weight and size. Greater hunting pressure is needed to suppress deer population growth in the study region, and continuous monitoring of captured deer is essential to evaluate population dynamics.
わが国においては、1980年代からニホンジカCervus nippon(以下、シカ)の個体数増加と分布拡大が進み、農林業被害をはじめとする経済被害や森林下層植生の衰退など生態系への影響が問題視されている(Kaji et al. 2022)。そのため、2013年度には環境省および農林水産省が抜本的な鳥獣捕獲強化対策を取りまとめ、2023年度までにシカ個体数を半減させるという目標を設定し、積極的な捕獲が推進された(環境省・農林水産省 2013)。しかしシカ個体数は依然として高水準にあり、また分布も拡大傾向が続いていることから(環境省 2021)、環境省および農林水産省は改めて2028年度までに個体数半減を目指す目標を再設定した(環境省・農林水産省 2023)。シカ個体群の動態は対象となる地域の気候、食物利用可能性、捕獲圧等様々な要因によって変動しうる(McCullough et al. 2009; Kaji et al. 2022)。個体群の齢構成や妊娠率といったパラメータは個体群の現在の状態を反映するとともに、個体群動態の予測精度を向上する上で重要な生態学的動態の過程に関する情報となる(飯島 2017)。そのため、対象地域に生息する個体群におけるこれらの基礎情報は、実態に即した適切な管理計画を策定する上で不可欠である。
浅間山は長野県と群馬県の県境に位置する活火山である。本山周辺に生息するシカは遺伝的に関東山地地域個体群の一部と考えられ(溝口・小西 2017)、1970年代後半には分布が確認されていなかったものの、2000年代に分布が拡大し個体数が増加したと考えられる(長野県 2001; 長野県 2021)。本地域では近年強い捕獲圧がかけられているにも関わらず依然として生息密度が高く、農林業被害や自然植生への影響が確認されている(長野県 2021)。そのため、シカによる被害を低減し、適正な生息密度に維持することを目的とした第二種特定鳥獣管理計画が策定されている(長野県 2021)。しかし本地域に生息するシカ集団について、齢構成や繁殖および成長特性といった個体群の質的評価および個体群動態予測の精度向上に必要な基礎情報は不足している。そこで本研究では、浅間山周辺地域で捕獲・収集されたシカ個体を対象に齢構成、繁殖および成長特性に関する基礎情報を明らかにすることを目的とした。
長野県側の浅間山周辺地域に生息するシカ集団を対象に、狩猟および許可捕獲により捕獲されたシカ個体の試料を収集した。本調査地域では捕獲方法としてわな猟が多く用いられている(小諸市野生鳥獣商品化施設、私信)。調査対象地域は図1の通りである。2013–2020年の各年12月下旬から5月上旬に捕獲された201頭(メス162頭、オス39頭)の頭部と子宮、および2023年7月に捕獲された100頭(メス50頭、オス50頭)の頭部、計301頭(メス212頭、オス89頭)の試料を収集した(付録1)。なお、オスの試料は2020年および2023年のみ収集した。

本調査地域は長野県が設定する管理ユニットのうち、越後・日光・三国管理ユニット、関東山地管理ユニット、八ヶ岳管理ユニットの3ユニットに属する(長野県 2021)。Iijima and Ueno(2016)に準拠したHarvest-basedモデルにより推定された生息密度は、2019年度末の時点で越後・日光・三国管理ユニットで9.27頭/km2、関東山地管理ユニットで31.42頭/km2、八ヶ岳管理ユニットで28.12頭/km2であり、3ユニットいずれの地域においても近年増加傾向を示している(長野県 2021)。これらの地域ではシカ個体数増加に伴い農林業被害や森林下層植生の衰退、高山植物の消滅といった問題が発生していることから、適正な生息密度維持が求められている(長野県 2021)。
齢構成本調査地域に生息するシカ集団の年齢分布を明らかにするために、各個体の年齢を以下の通り査定した。2013–2020年に捕獲された個体については下顎骨の歯の萌出・交換状況(大泰司 1980)により0歳と1歳以上の個体に分類した。2013–2020年に捕獲され1歳以上と分類した個体および2023年に捕獲されたすべての個体の下顎骨から第一切歯を収集し、セメント質年輪法(大泰司 1980; Hamlin et al. 2000)により捕獲時の年齢を査定した。なお、セメント質年輪法に基づく齢査定は全てMatson’s lab(Manhattan, Montana, USA)により行われた。査定された齢情報を基に、各齢の個体数を集計した。日本に生息するニホンジカを対象とした多くの先行研究では、2歳以上の個体を成獣として区分している(横山ほか 2003; Ueno et al. 2018; 松金・横山 2018)。そのため本研究においても、0歳および1歳を「若齢個体」、2歳以上を「成獣個体」と区分し、各区分の個体数に着目した。
繁殖特性本調査地域に生息するシカ集団の繁殖特性を明らかにするために、2013–2020年の各年12月下旬から5月上旬に捕獲されたメス個体を対象に以下項目を算出した。
齢別妊娠率を算出するために、2013–2016年に捕獲された個体については捕獲時の子宮内の胎子の有無に関する記録をもとに、2018–2020年に捕獲された個体については著者らの直接観察に基づく子宮内の胎子の有無により妊娠判定を行った。妊娠判定できた160頭を対象に、齢階級ごとに妊娠率を算出した。なお、日本に生息する二ホンジカは通常1歳から繁殖活動を開始することが知られており(Ohnishi et al. 2009)、本調査地域においても同様であることが確認されている(Hata et al. 2021)。そのため本論文では1歳を性成熟齢として取り扱うことにした。
ニホンジカと近縁であるアカシカCervus elaphusにおいては、個体群密度の増加等に伴い母親の栄養ストレスが上昇するとオスが産まれる割合が減少することが報告されている(Kruuk et al. 1999; Mysterud et al. 2000)。そこで本調査地域において胎子性別に偏りがあるかどうかを検討した。2018–2020年に収集した試料からは胎子を得ることができたため、胎子の性別を外部生殖器の形態から判断し、各性別の個体数を計数した。続いて目的変数を胎子数、説明変数を性別とした一般化線形モデルを構築し、性比の偏りが生じているかを検討した。なお、年によって収集サンプル数にばらつきがあるため、offset項として捕獲年を設定した。
シカ類において繁殖可能となる体重には閾値が存在することが報告されている(Solberg et al. 2002; Miura and Tokida 2009; Flajšman et al. 2017)。本集団において性成熟齢の個体は妊娠有無に応じて体重に差があるかを明らかにするために、体重データが得られた2018–2020年に捕獲された性成熟齢のメス個体を対象に、Mann–WhitneyのU検定により妊娠個体と非妊娠個体の体重を比較検討した。なお、妊娠週数の差に伴い体重のばらつきが発生することを考慮して、妊娠中期にあたる1–3月に捕獲された性成熟齢個体のみを対象に解析を行った(n=17)。
最後に受胎日および出産日を推定するために、2018–2020年に捕獲された各胎子の胎齢を推定した。なお胎子の胎齢推定を行う場合、ニホンジカの体サイズ(出生時体重)は地域個体群によって変異が大きいことに留意する必要があるため(Suzuki et al. 1996)、本研究では本調査地域と地理的に比較的近接した栃木県のシカ個体群を対象に作成された胎齢推定式(岩本ほか 2009)を使用した。胎齢が推定できた胎子を対象に(n=45)、推定受胎日を算出した。また、妊娠期間を231日として(岩本ほか 2009)、推定受胎日から起算した推定出産日を算出した。
成長特性体サイズは食物の利用可能量や質によって左右されることが知られている(Huston and Wolverton 2011)。本調査地域に生息するシカ集団の体重および体サイズの成長特性を明らかにするために、2018–2020年および2023年に収集した個体を対象として、体重(n=221)および体サイズ指標である頭蓋骨最大長(Total Length、以下TL; von den Driesch 1976; Hata et al. 2021)(n=222)の成長曲線の推定を行った。まず体重データとして、捕獲時に測定された体重を使用した。続いてTLデータとして各個体のTLを計測した。計測はデジタルノギスを用いて0.01 mm単位で測定した。
得られたデータをもとに成長曲線を推定するにあたり、実データに対してより当てはまりの良いモデルを選択するために、3モデル(ベルタランフィ—の成長曲線・ロジスティック曲線・ゴンペルツ曲線)を当てはめた。各モデルは以下の通り記述される。
 | (式1) |
 | (式2) |
 | (式3) |
tは月齢を示し、L(t)はt時点の体重あるいはTLを示す。aは漸近値(最大到達体重あるいは最大到達TL)、bは時間スケールに関するパラメータ、eは自然対数の底、kは成長係数、t0は体重あるいはTLがゼロとなる時の月齢を示す(Eisen et al. 1969; Bartareau et al. 2011)。pは体重の成長曲線には3を、TLの成長曲線には1をそれぞれ代入した(Bartareau et al. 2011)。月齢は齢査定結果をもとに、6月1日を出産日と仮定して算出した。月齢の算出にあたっては日本に生息するニホンジカを対象とした多くの既往研究で6月1日が起算日として採用されていること(例えばSuzuki et al. 2001; 横山ほか 2003)、本地域の推定出産日が5月下旬をピークとすることからも(結果参照)、6月1日を起算日とすることが妥当であると考えた。なお、本研究で得られたシカ試料は2018–2020年12月下旬から5月上旬および2023年7月に捕獲されており、季節による体重変動が懸念された。体重の成長曲線を推定する場合、季節による体重変動幅の大きさを推定するために正弦あるいは余弦曲線を当てはめる場合がある(Leader-Williams and Ricketts 1981; Suzuki et al. 2001)。本研究では季節による変動幅そのものよりも、季節による変動を内包した上で漸近値に到達する年齢および体重値に着目するために、正弦あるいは余弦曲線を当てはめずに推定を行った。各パラメータ値は関数nls()を用いて推定を行った。なお、ゴンペルツ曲線の推定には関数SSgompertz()も併せて使用した。各モデルの当てはまりの程度を検討するために赤池情報量基準(AIC)を算出し、AICが最も小さい成長曲線モデルを採用した。採用された成長曲線モデルに基づき、体重あるいはTLの漸近値の98%に達する月齢を算出し、年齢に換算した。全ての統計解析はR for Windows 4.2.2(R Development Core Team 2012)で行った。
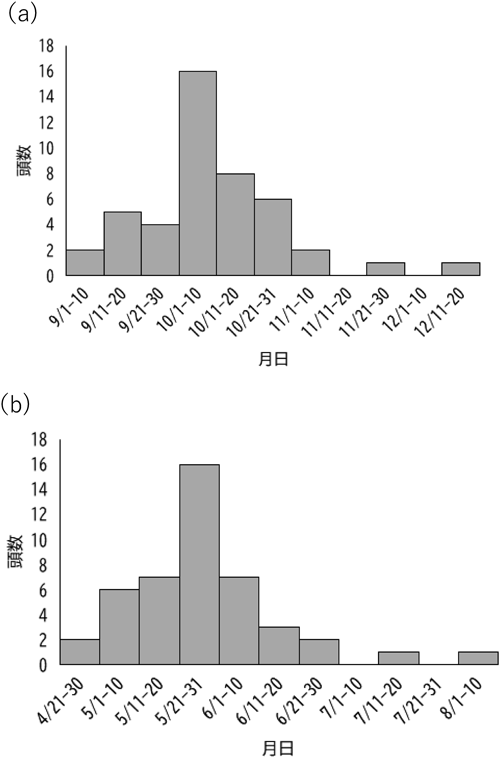
2013–2020年および2023年に本調査地域で捕獲されたシカのうち、齢査定できた298頭(メス210頭、オス88頭)の年齢ごとの個体数を図2に示す。捕獲した季節の違いが年齢分布に影響を及ぼすと考えられたことから、2013–2020年各年12月下旬から5月上旬に捕獲・収集された個体(図2a)と2023年7月に捕獲・収集された個体(図2b)でそれぞれ集計した。若齢個体(0–1歳)が全体に占めた割合は2013–2020年が43.4%、2023年が54.0%であった。メスを対象に比較的サンプル数が多かった2018–2020年および2023年の若齢個体割合の経年変化を調べた結果、2018年36.0%(n=50)、2019年36.4%(n=44)、2020年51.7%(n=29)、2023年48%(n=50)であった。なお、捕獲個体のうち、メスの最高齢は18歳、オスの最高齢は12歳であった。

2013–2020年に捕獲されたメス160頭を対象に平均妊娠率を齢別に算出した結果を図3に示す。0歳での妊娠例が1例のみ認められた。性成熟齢(1歳)での妊娠率は50.8%、成獣個体(2歳以上)の妊娠率は91.0%であった。なお、サンプル数がある程度確保できた年(n≧20)に絞って成獣個体の妊娠率の経時変化を検討した結果、2018年87.5%(n=50)、2019年100%(n=44)、2020年85.7%(n=29)であった。高齢になると妊娠率は低下し、10歳以上個体の妊娠率は76.9%であった。14歳の捕獲個体は本研究では得られず、確認できた妊娠個体の最高年齢は13歳であった。なお、15歳以上で妊娠した個体は認められなかった。

2018–2020年に捕獲されたメスジカの子宮内から摘出した胎子のうち、性別が確認できた74頭を対象として性別ごとの個体数を集計した(図4)。性比はオス : メスが2018年1 : 0.72、2019年1 : 1.08、2020年1 : 0.78となった。一般化線形モデルによる解析の結果、性比に有意な偏りは認められなかった(P=0.66)。

2018–2020年に捕獲されたメス123頭の年齢と体重の関係を図5に示す。各齢の妊娠個体の体重には大きなばらつきがみられた。妊娠中期にあたる1–3月に捕獲された性成熟齢個体において、妊娠個体(n=9)の平均体重は46.7 kg(sd=7.2)であり、非妊娠個体(n=8)の平均体重は33.8 kg(sd=2.7)と有意に低かった(P<0.01)。なお、性成熟齢で妊娠が確認された個体の最低体重は37.68 kgであった。

推定された受胎日は9月上旬から12月中旬にかけて分布し、10月上旬がピークとなった(図6a)。また、推定出産日は4月下旬から8月上旬にかけて分布し、5月下旬がピークとなった(図6b)。
体重およびTLの成長曲線についてデータへの当てはまりの良さを検討するために3モデルの各パラメータ値を推定したところ、メスの体重成長曲線のみゴンペルツ曲線モデルが最もAIC値が低かったものの、オスの体重成長曲線および両性のTL成長曲線はロジスティック曲線モデルが最もAIC値が低くなった(表1)。メスの体重成長曲線についても、ゴンペルツ曲線モデルとロジスティック曲線モデルのAIC値差は0.39と僅かであったため、本研究では成長曲線モデルとしてロジスティック曲線モデルを採用した(表1)。ロジスティック曲線モデルによって推定された体重およびTL成長曲線を図7に、各パラメータ値を表1に示す。なお、本研究で得られた実際の最大体重は、メスは72.44 kg(8歳)、オスは96.76 kg(9歳)であった。また実際の最大TLは、メスは282.72 mm(3歳)、オスは310.98 mm(9歳)であった。体重の漸近値(α)の98%に到達する年齢を算出した結果、メスは5.31歳、オスは12.28歳と推定された。TLの漸近値(α)の98%に到達する年齢を算出した結果、メスは3.51歳、オスは7.52歳と推定された。
| (a)メスモデル | |||||||
|---|---|---|---|---|---|---|---|
| 目的変数 | 供試数 | モデル | α | b | k | t0 | AIC |
| 体重(kg) | 144 | ベルタランフィ成長曲線 | 57.38(0.95) | — | 0.05(0.01) | −20.03(3.53) | 922.47 |
| ロジスティック曲線 | 57.06(0.89) | 1.84(0.21) | 0.07(0.01) | — | 922.76 | ||
| ゴンペルツ曲線 | 57.30(0.93) | 1.17(0.10) | 0.94(0.01) | — | 922.37 | ||
| 頭蓋骨最大長(TL、mm) | 145 | ベルタランフィ成長曲線 | 269.84(1.19) | — | 0.06(0.01) | −16.64(2.16) | 1024.63 |
| ロジスティック曲線 | 269.53(1.15) | 0.46(0.03) | 0.07(0.01) | — | 1024.21 | ||
| ゴンペルツ曲線 | 269.68(1.17) | 0.40(0.03) | 0.93(0.01) | — | 1024.34 | ||
| (b)オスモデル | |||||||
| 目的変数 | 供試数 | モデル | α | b | k | t0 | AIC |
| 体重(kg) | 77 | ベルタランフィ成長曲線 | 99.30(8.10) | — | 0.02(0.00) | −3.57(0.59) | 547.57 |
| –51.28(13.53) | |||||||
| ロジスティック曲線 | 92.88(4.76) | 2.69(0.32) | 0.03(0.01) | — | 544.51 | ||
| ゴンペルツ曲線 | 96.89(6.78) | 1.41(0.10) | 0.98(0.00) | — | 546.73 | ||
| 頭蓋骨最大長(TL、mm) | 77 | ベルタランフィ成長曲線 | 296.69(7.79) | — | 0.03(0.01) | −42.93(12.30) | 637.95 |
| ロジスティック曲線 | 295.47(7.02) | 0.42(0.05) | 0.03(0.01) | — | 637.73 | ||
| ゴンペルツ曲線 | 296.02(7.36) | 0.36(0.04) | 0.97(0.01) | — | 637.84 | ||

シカ個体群は対象となる地域の気候、食物利用可能性、捕獲圧等様々な要因によって変動しうる(McCullough et al. 2009; Kaji et al. 2022)。全国的に個体数が増加し分布が拡大傾向であるシカ管理において、各地域に応じたシカ個体群の性状把握は、個体群動態の予測精度を向上させ、より適切な管理計画を策定するための重要な情報源となる。本研究では、近年生息密度が増加傾向にある浅間山周辺地域のシカ個体群の質的評価に資する齢構成や繁殖・成長特性といった基礎情報を明らかにした。
齢構成本調査地域で捕獲されたシカの齢構成において若齢個体(0–1歳)が占める割合は、調査期間中経年的に増加傾向を示した。ただし2023年は0歳捕獲個体が0頭であったことに留意する必要がある。2023年に捕獲された個体に0歳が含まれていないのは、7月に捕獲された個体を収集したためであると考えられる。先行研究から本州におけるシカの出産時期は5–6月にピークを迎え、8–9月まで及ぶ場合があることから(岩本ほか 2009; Koizumi et al. 2009)、7月の時点では未出生あるいは出生直後のため行動を開始していない可能性がある。また出生後であっても夏季の幼獣は休息行動の占める割合が多い(相馬ほか 1994)。そのため、7月に0歳個体を捕獲する機会は低いと考えられた。0歳不在の状態においても2023年は若齢個体割合がメスで48.0%であったことから、齢構成に占める若齢個体割合は2018年(36.0%)や2019年(36.4%)と比較して高いと推察される。小泉(2006)は狩猟者が0歳を射殺することを忌避する傾向にあることから、捕獲圧の増加に伴い捕獲個体の齢構成が若齢化することを指摘している。しかしながら本調査地域では捕獲方法としてわな猟が多く用いられていることから(小諸市野生鳥獣商品化施設、私信)、捕獲圧が齢によって大きく異なるとは考えにくい。一般的に有蹄類の個体群では密度が増加すると若齢個体の死亡率が高くなり、妊娠率が低下することが知られている(Gaillard et al. 1998; Gaillard et al. 2000)。本調査地域では生息密度が増加傾向であること(長野県 2021)を考慮すると、メスの若齢個体割合が経年的に増加しているという結果は、密度増加に伴う若齢個体の生存率の低下等が生じておらず、集団が依然として高い繁殖力を維持している状態であることを示唆している。
繁殖特性0歳における妊娠例はごく少数ではあるが報告されており(飯島 2011; Matsuura 2022)、非常に稀ではあるものの発生しうるものと考えられた。性成熟齢(1歳)での妊娠率は栄養状態によって大きく変動することが知られており、兵庫県では密度増加に伴い1988–1992年(平均生息密度12.8頭/km2; Koizumi et al. 2009)は76.6%だった妊娠率が、2003–2015年(平均生息密度20–30頭/km2)には15.6%まで減少した(松金・横山 2018)。他地域における性成熟齢での妊娠率は山梨県(平均生息密度33.3頭/km2;山梨県 2022)で77%(飯島 2011)、熊本県(平均生息密度19.2頭/km2)で84%(Koizumi et al. 2009)との報告があり、他地域と比較すると本調査地域(平均生息密度9.27~31.42頭/km2)の妊娠率(50.8%)はやや低いと考えられた。一方で本調査地域の成獣個体(2歳以上)の妊娠率は91.0%であり、2018–2020年の3年間の経年変化も85.7–100%と高値を推移していた。高栄養な環境下では成獣メスの妊娠率は80%を超えることが報告されている(Ohnishi et al. 2009)。密度増加に伴う若齢個体の妊娠率変動が生じた兵庫県においても2歳以上の妊娠率は比較的安定していた(1988–1992年:89.4%、2003–2015年:80%以上)(松金・横山 2018)。また、他地域においても2歳以上の妊娠率は山梨県(平均生息密度33.3頭/km2)で77%(飯島 2011)、熊本県(平均生息密度19.2頭/km2)で2歳93%および3歳以上92%(Koizumi et al. 2009)と高い値であることが報告されている。そのため、本調査地域は他地域と同様、成獣が高い妊娠率を維持する程度の食物資源が依然として存在していると考えられた。特に本調査地域には牧草地やゴルフ場といった人工草地が多く点在し、人工草地率が高い場所は個体数増加率および環境収容力が高くなることが報告されていることから(Iijima and Ueno 2016)、こうした場所が安定した食物供給源としての機能を果たしていると推測される(Hata et al. 2019)。高齢個体の妊娠率に着目すると、10歳以上個体の妊娠率は76.9%であった。15歳以上で妊娠した個体はおらず、妊娠最高齢は13歳であった。10歳以上個体の妊娠率は兵庫県(平均生息密度12.8頭/km2)で86.6%、熊本県(平均生息密度19.2頭/km2)で92%との報告があり(Koizumi et al. 2009)、これらの地域と比較するとやや低い結果となったが、高齢個体のサンプル数の少なさも影響したと考えられた。なお、子宮内胎子を肉眼的に観察できるのは胎齢30日頃とされ(鈴木 1994)、本調査地域における推定受胎日が12月中旬まで及ぶことから、12月下旬から1月中旬頃に捕獲された個体の妊娠率を過小評価した恐れがある。しかしながら妊娠判定を行った160頭のうち当該期間に捕獲された個体は19頭であり、このうち1歳以上で妊娠が確認された個体の割合は80%であったことから、採材時期が本研究で算出した妊娠率にもたらした影響は大きくないと考えられた。
胎子性比(オス : メス)はこれまでにも北海道、兵庫県、岩手県金華山島に生息するシカ個体群を対象に検討され、いずれも有意な差は認められていない(Ohnishi et al. 2009; 松金・横山 2018)。アカシカでは母親の栄養状態と出生オス数は相関し、個体数の増加や気候の厳しさに起因する栄養ストレスが増加すると出生オス割合が減少することが報告されている(Kruuk et al. 1999; Mysterud et al. 2000)。本調査地域においても日本国内の他地域と同様に性比に有意な差が認められなかったことから、繁殖活動に参加するメスの栄養状態は出生子の性比に偏りをもたらすほど悪い状況ではないと考えられた。
シカ類において繁殖可能となる体重には閾値が存在することが報告されている(Solberg et al. 2002; Miura and Tokida 2009; Flajšman et al. 2017)。他地域において繁殖可能となる閾値体重は北海道中島で43 kg、宮城県金華山島で40 kgと報告されている(Miura and Tokida 2009)。本調査地域では、繁殖活動を開始すると考えられる1歳妊娠個体の体重は1歳非妊娠個体よりも有意に重かった。1歳妊娠個体の平均体重が46.7 kgであったこと、1歳妊娠個体のうち最低体重は37.68 kgであったことから、本調査地域では北海道や宮城県の個体群と同じく概ね40 kg前後が繁殖活動開始の閾値体重となると考えられた。また、本調査地域において2歳以上では1歳妊娠個体の平均体重以下の個体でも妊娠している個体がみられた(図5)。これは、加齢に伴い体重が妊娠率にもたらす影響が小さくなったため(Borowik et al. 2016; Flajšman et al. 2017)と考えられた。
本調査地域の受胎日は9–12月でピークが10月、出産日は4–8月でピークが5月であった。他地域における受胎日および出産日については、北海道で受胎日が10–1月でピークが10月、出産日のピークが6月(Suzuki et al. 1996; Matsuura 2022)、栃木県で受胎日が9–12月でピークが10月、出産日が5–8月でピークが5月下旬(岩本ほか 2009)、兵庫県で受胎日が8–1月でピークが9月、出産日が4–9月でピークが5月(Koizumi et al. 2009)であったことが報告されている。これらの報告と比較すると、本調査地域の受胎日および出産日は地理的に比較的近い栃木県の個体群とほぼ同時期であると推定された。
成長特性体重の成長曲線を推定し、漸近値(α)の98%に到達する年齢を算出した結果、メスは約5歳まで、オスは約12歳まで増加すると考えられた。また、最大体重はメスが72.44 kg、オスは96.76 kgであった。Suzuki et al.(2001)は北海道東部(斜里町・足寄町・音別町)で 1987–1993年に捕獲されたシカ(1993年足寄町の平均生息密度2.3頭/km2;梶ほか 2006)を対象に、体重成長曲線がメスは3–4歳頃に、オスは6–7歳頃に平衡に達することを報告している。またYokoyama(2009)は兵庫県で2002–2007年に捕獲されたシカ(2002年の平均生息密度約20頭/km2;横山・松金 2018)を対象に、体重成長曲線がメスは3歳まで、オスは5歳まで増加することを報告している。最大体重は北海道東部でメスが96 kg、オスが150 kg(Suzuki et al. 2001)、兵庫県でメスが約60 kg、オスが約85 kgであった(Yokoyama 2009)。日本においてはシカの体サイズが緯度の上昇に伴い大きくなる傾向が認められており(Kubo and Takatsuki 2015)、本調査地域は上述した2調査地の中間にあたると考えられる。最大体重は他地域と比較して特異的な値ではないものの、体重成長が頭打ちとなる年齢は両地域よりも特にオスで高い傾向にあった。
続いて体サイズの指標として頭蓋骨最大長(TL)の成長曲線を推定した結果、メスは約3–4歳、オスは約7–8歳頃までに成長が概ね完了すると考えられた。体重成長曲線はメスで約5歳まで、オスで約12歳まで増加すると推定されたことから、TLの成長が完了したあとも、体重はより長い期間増加し続けると考えられた。体サイズ指標と体重の成長時期のずれは他地域でもみられ(Kaji et al. 1988; Yokoyama 2009)、成長ピークが筋肉や脂肪組織と比較して骨の方が早く訪れることが原因と考えられる(Reimers 1972)。他地域における体サイズ指標の成長速度を検証した事例として、北海道東部でメスが肩高3歳、体長3歳、後足長2歳、オスが肩高3歳、体長4歳、後足長2歳で平衡に達することが報告されている(Suzuki et al. 2001)。また兵庫県で体高および後足長が両性共に2歳で成長が完了することが報告されている(Yokoyama 2009)。いずれの先行研究も本研究とは異なる体サイズ指標を用いているものの、これらと比較すると本調査地域のシカ個体群は個体の成長が遅れており、最大値に到達するまでの時間が長いと考えられた。
動物の体サイズは食物の利用可能性と質によって左右されることが知られている(Huston and Wolverton 2011)。他地域と比較して、本調査地域に生息するシカ集団がより長い時間をかけて体重やTLが最大値に到達する理由として、高密度化に伴う食物制限による影響が考えられる。先行研究において、ノロジカCapreolus capreolusでは体サイズおよびその成長速度が密度に関係し、低密度環境で早く大きく成長することが示されている(Klein and Strandgaard 1972)。また高密度環境下に生息するオジロジカOdocoileus virginianusは、体サイズおよびその成長速度を調整することで高密度状態を維持することが報告されている(Simard et al. 2008; Wolverton et al. 2009)。本調査地域では上述の通り、依然として成獣メスが高い妊娠率を維持する程度には食物資源が存在すると考えられる一方で、高密度化に伴い体重や体サイズの成長速度の低下という形質変化が進行しているのかもしれない。また、最大値への到達年齢が遅い傾向がオスでより顕著であった理由として、シカ類ではオスの方がメスより栄養要求量が高く(Rose et al. 1998)、体サイズ成長において食物条件の影響をより強く受けることが関係していると考えられた(Clutton-Brock et al. 1982; Kaji et al. 1988)。
まとめ浅間山周辺地域に生息するシカ集団は、メスの齢構成に占める若齢個体割合が増加傾向にあること、メス成獣が高い妊娠率を維持していたことから、依然として高い繁殖力を維持している可能性が高いと考えられた。一方で繁殖活動を開始する年齢と考えられる1歳妊娠率が他地域と比較してやや低いこと、体重およびTLの成長に年数を要していることから、高密度化に伴う影響が生じている可能性が考えられた。Eberhardt(2002)は密度上昇に伴い低下する生活史形質には順序が存在することを示しており、その順序は幼獣生存率の低下、初産齢の遅延、成獣妊娠率の低下、そして成獣生存率の低下であるとしている。本研究では幼獣生存率を直接検証することができなかったものの、メスの齢構成に占める若齢個体割合が経年的に増加したことから、幼獣生存率の低下が生じている可能性は低い。一方で密度増加に伴い次に生じるとされる初産齢の遅延については、繁殖活動を開始すると考えられる1歳妊娠率(50.8%)が他地域と比較してやや低い傾向であったことから(Koizumi et al. 2009; 飯島 2011; 松金・横山 2018)、徐々に進行しはじめている可能性があると考えられた。しかし、今後本調査地域において高密度化に伴う自然な個体数減少が生じることは期待できない可能性が高い。なぜなら密度効果によって個体群成長が制限されることがあったとしても、個体群調節と呼べる効果が観測された事例は非常にわずかであり(上野 2017)、上位捕食者が不在な環境に生息するシカ類の個体数と密度を制限する主要因は捕獲圧であるとされているからである(Rooney and Waller 2003; Ueno et al. 2010)。対象地域における集団の個体数増加を抑制するためには、引き続き高い捕獲圧をかけ続ける必要があるとともに、捕獲個体の分析によるモニタリングを継続して行い、集団の各パラメータ値の変化を評価し、管理方針に還元することが重要である。
捕獲シカ試料の収集にご協力いただいた狩猟者および小諸市野生鳥獣商品化施設のみなさま、竹下 毅氏に感謝します。本研究で収集した試料の多くは小諸市野生鳥獣商品化事業のもと収集されました。試料処理にあたっては、小山彩由里氏、真中幸子氏、農研機構・畜産研究部門の坂本ゆかり氏およびつくば第1業務科の荻津 勝氏・小川鷹祐氏、日本獣医生命科学大学の嶌本 樹氏・飯沼凛湖氏・塚田栞子氏の協力をいただきました。本研究はJSPS科研費JP19K20492・21H03658・23K17072の助成を受けたものです。
付録1 表1. 本研究で使用したシカ標本一覧。
各個体の属性情報を示す。各項目のデータは本文中に記載の方法で取得した。なお、NAはデータが得られなかったことを示す。
リンクが示されていない付録は本文のオンラインサイトに掲載。
https://doi.org/10.18960/hozen.2425
