‘メロン中間母本農 5 号’ は,固定品種 ‘アールスフェボリット春系 3 号’ (‘ 春系3 号’)にメロン退緑黄化病抵抗性を有する系統 I-10 を交雑し,抵抗性検定による選抜と自殖を繰りかえすことにより,メロン退緑黄化病抵抗性を固定し,雌花着生性等の主要形質を向上させた中間母本である.本品種はメロン退緑黄化病に抵抗性を有する.高温等の条件で発病する場合があるが,発病した場合でも罹病性品種に比べて発病程度はきわめて軽く,ウリ類退緑黄化ウイルスの感染が本品種の果実品質に与える影響はほとんど認められない.メロン退緑黄化病抵抗性は劣性に遺伝し,1 つの主要な遺伝子が関与すると推察され,SSR マーカー ECM230 は,メロン退緑黄化病抵抗性育種における選抜マーカーとして利用できる.本品種の節間および葉柄は長く,雌花着生率は素材である I-10 に比べ高い.成熟日数は 45 日程度で,‘春系 3 号’ に比べ 10 日ほど早く,離層が形成されるためへた離れしやすい.果実は偏平で,‘春系 3 号’ に比べ小さく,溝があり,花痕部は大きい.糖度は I-10 に比べ高く,11° Brix 程度である.
メロン退緑黄化病は,クリニウイルス属に属するウリ類退緑黄化ウイルス(Cucurbit chlorotic yellows virus,CCYV)を病原とするウイルス病である( 行徳ら 2009,Okuda et al. 2010).CCYV は,2004 年に熊本県のメロン産地で初めて発生が報告された.現在では,九州全域,中国,四国,近畿,東海,関東地域のウリ科産地で発生し問題となっている.海外では,中国(Gu et al. 2011),台湾(Huang et al. 2010),スーダン(Hamed et al. 2011),エジプト(Amer 2015),レバノン(Abrahamian et al. 2012),サウジアラビア(Al-Saleh et al. 2015),イラン(Bananej et al. 2013),ギリシャおよびトルコ(Orfanidou et al. 2014,2017),アメリカ合衆国(Wintermantel et al. 2019)での発生が報告されている.CCYV は,タバココナジラミバイオタイプ Q(Bemisia tabaci Mediterranean(Q biotype))およびバイオタイプ B(B. tabaci Middle East–Asia Minor 1(B biotype))によって半永続的に伝搬され(行徳ら 2009),本ウイルスに感染した植物は,葉に退緑小斑点を生じ,その後葉全体が黄化する.果実に病斑の出現は認められないが,メロン(Cucumis melo L.)では果実重および糖度の低下が問題となり,特にアールス系メロンの抑制栽培での被害が深刻である.メロン退緑黄化病の防除には媒介虫であるタバココナジラミを駆除することが重要であるが,タバココナジラミバイオタイプ Q は一部のネオニコチノイド系薬剤に対する感受性が低く,それを完全に防除することは容易ではない.生産者からは抵抗性品種の導入が強く求められているが,これまで抵抗性品種および素材の報告はなかった.そこで,筆者らは,メロン退緑黄化病抵抗性メロンの検索を行い,インド原産の I-10(JP 138332)が抵抗性を有することを明らかにし(Okuda et al. 2013),I-10 を素材としてメロン退緑黄化病に抵抗性を有する‘ メロン中間母本農 5 号’を育成した.ここにその育成経過と特性について報告する.
2009 年から I-10 を素材としてメロン退緑黄化病抵抗性メロンの育成を開始した.I-10 は,果皮が白色で緑色の斑紋があり,雌花着生が少ないことや,果肉糖度が市販品種(13 ~ 14° Brix)に比べきわめて低い(8° Brix 程度)こと等の欠点を有する.そこで,I-10 にわが国の固定品種 ‘ アールスフェボリット春系 3 号’(‘春系 3 号’)を交雑し,抵抗性検定による選抜・固定を進めた.2014 年に育種素材の I-10 に比べ,雌花着生率が高く,果皮および果肉色が淡緑で,果肉糖度が 11° Brix 程度に向上した系統を得た.本系統はメロン退緑黄化病に対して I-10 と同様に抵抗性を有することが判明したことから,本系統にメロン久安 1 号の系統名を付与した(図 1).2015 および 2017 年度にメロン退緑黄化病抵抗性検定試験を実施したところ,抵抗性が確認された.次に,罹病性のアールス系メロン ‘AnMP-5’ とメロン久安 1 号との交雑後代を用いてメロン退緑黄化病抵抗性の遺伝性を調査した結果,抵抗性は劣性に遺伝し,1 つの主要な遺伝子が関与すると推定された.これらのことから,メロン久安 1 号は中間母本として有望であると判断し,2019年に ‘メロン中間母本農 5 号’ として品種登録出願した(品種登録出願番号第 33973 号,2019 年 6 月 6 日).

1 試験成績
試験の概要を表 1 に示す.

a メロン退緑黄化病抵抗性
メロン退緑黄化病抵抗性検定は,2015 および 2017 年度に九州沖縄農業研究センター筑後・久留米研究拠点(福岡県久留米市)で実施した.対照品種として,罹病性の ‘ミラノ夏Ⅰ’ および抵抗性の I-10 を用いた.CCYV 感染キュウリを餌としてタバココナジラミバイオタイプ Q を飼育している網室にメロン苗を 10 ~ 14 日間置くことで CCYV の接種を行った.接種後,メロン苗に付着しているタバココナジラミを薬剤により殺虫し,ビニールハウス内に定植した.罹病性品種の本葉に病徴がはっきりと確認された接種 2 か月後以降に,主枝の第 5 ~ 14 本葉の発病評点(0:無病徴,1:退緑斑が確認できる,2:黄化が確認される(全体の1 ~ 20%),3:黄化が広がり,斑状となる(全体の 20 ~ 50%),4:黄化が全体に広がる(全体の 50%以上))から平均発病評点を算出し,抵抗性を判定した.
2015 年度の試験では,‘ メロン中間母本農 5 号’および I-10 における全ての株で発病せず,病徴は認められなかった(図 2,表 2).一方,2017 年度の試験では,‘メロン中間母本農 5 号’ および I-10 の多くの個体で弱い病徴が認められ,発病評点は,それぞれ 1.2 および 2.1 であった.‘ミラノ夏Ⅰ’では両試験の全ての株で発病評点が 3.5 ~ 4.0 の激しい黄化症状が認められた.‘メロン中間母本農 5 号’ は,発病した場合でも病徴は罹病性品種に比べ非常に弱く,メロン退緑黄化病に対して実用的に利用可能な抵抗性を有すると考えられた.


b CCYV の感染が収量および果実品質に与える影響
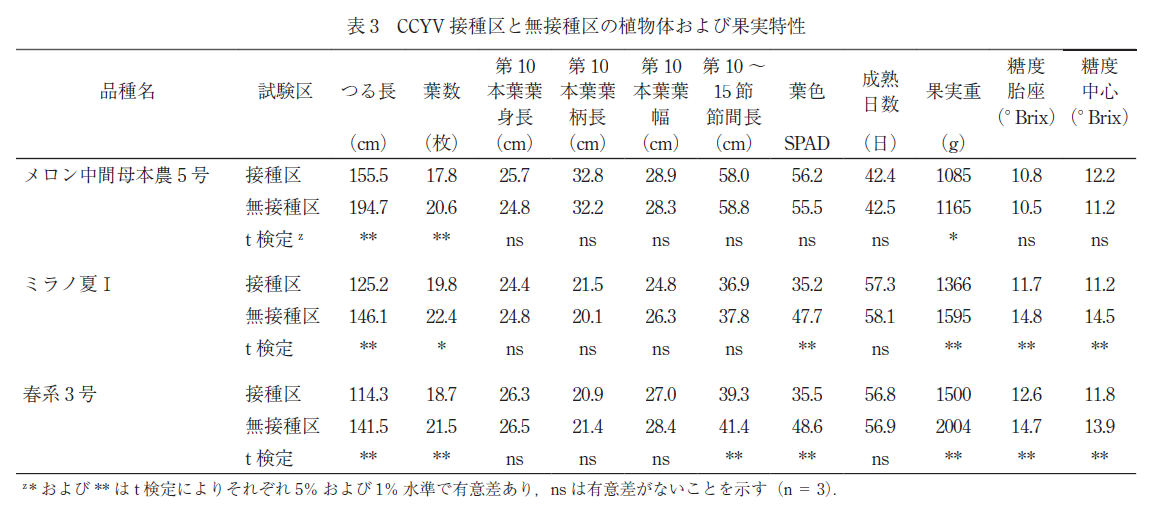
CCYV の感染が収量および果実品質に与える影響の評価は,2014 年度に九州沖縄農業研究センター筑後・久留米研究拠点で実施した.‘ メロン中間母本農 5 号’,罹病性の ‘ミラノ夏Ⅰ’ および ‘春系 3 号’ を用いた.
CCYV の接種は上記のメロン退緑黄化病抵抗性の評価と同様に行い,CCYV を接種した接種区と無接種区を設定した.接種後の植物をビニールハウス内に定植し,主枝は畝面から約 180 cm の高さにある誘引線に達した段階で摘心した.主枝第 9 節までに発生した側枝を摘除し,第 10 ~ 13 節から発生した側枝第1 節に着生した雌花を目標に交配を行い,1 株に 1 果を着果させた.接種区および無接種区の植物体の生育,ならびに果実品質を調査し,CCYV 感染の影響を調査した.
全品種の接種区におけるつる長および葉数は無接種区に比べ,低い値となった(表 3).この原因として,無接種区の育苗ではタバココナジラミを放飼しておらず,コナジラミの吸汁害がなかったのに対し,接種区の育苗では 1 株あたり 100 頭以上のコナジラミを接種したため,この吸汁害によって初期生育が大きく阻害されたことが考えられた.罹病性品種である ‘ミラノ夏I’ と ‘春系 3 号’ の接種区の SPAD 値は,それぞれ 35.2 および 35.5 で,無接種区の 47.7 および 48.6 に比べ有意に低くなった.一方,‘ メロン中間母本農5 号’では,接種区で 56.2,無接種区で 55.5 となり,両区の有意差は認められなかった.‘ メロン中間母本農 5 号’ では,接種区の果実重は 1085 g で,無接種区の 1165 g よりやや軽かったが,果肉の胎座部および中心部の糖度に差は認められなかった.罹病性品種では,接種区における果実重は軽く,‘春系 3 号’ および ‘ミラノ夏Ⅰ’ の果肉糖度(胎座)は,それぞれ 11.7 および 12.6° Brix で,無接種区に比べ有意に低くなった.
c メロン退緑黄化病抵抗性の遺伝解析
メロン退緑黄化病抵抗性の遺伝様式の解析は,2015 年度に九州沖縄農業研究センター筑後・久留米研究拠点で実施した.罹病性 ‘AnMP-5’ と ‘ メロン中間母本農 5 号’ との F2 集団 114 個体,各 10 個体の親品種,F1を用いた.CCYV の接種および抵抗性の判定は,上記のメロン退緑黄化病抵抗性評価と同様に行った.また,Kawazu et al.(2018)は,‘AnMP-5’ と I-10 の F2 集団を用いて QTL 解析を行い,I-10 由来の 1 つの主要な QTL が第 1 染色体に座乗することを報告している.そこで,‘AnMP-5’ と ‘ メロン中間母本農 5 号’ とのF2 集団において,I-10 由来の QTL 近傍の SSR マーカー ECM230 を用いて,マーカー遺伝子型と発病評点との関連性を調べた.各個体のマーカー遺伝子型を調べるため,まず,DNeasy 96 Plant Kit(QIAGEN) を用いて, 各個体の本葉からゲノム DNA を抽出した.次に,Shimizu et al.(2011)が報告した 1 チューブ多重ポストラベル法を用いて,PCR 産物を蛍光色素標識し,シーケンサー 3730xl DNA Analyzer(Applied Biosystems) で泳動し,GeneMapper(Applied Biosystems)ソフトウェアで PCR 産物の長さを解析した.PCR の温度条件は,95 ℃ 5 分間,「95 ℃ 20 秒間,55 ℃ 90 秒間,7 2℃ 30 秒間」を 33 サイクル,「95 ℃ 20 秒間,49 ℃ 90 秒間,72 ℃ 30 秒間」を 3 サイクル,72 ℃ 5 分間とし,酵素は QIA GEN Multiplex PCR Kit(QIAGEN)を用いた.
‘メロン中間母本農 5 号’ と ‘AnMP-5’ との交雑 F1 の発病評点は,‘AnMP-5’ の評点 3.5 とほぼ同じ 3.2 および 3.6 であった(表 4).このことから,‘ メロン中間母本農 5 号’が有するメロン退緑黄化病抵抗性は劣性に遺伝することが明らかになった.交雑 F2 集団の発病評点は 0 ~ 4.0 の分布を示し(図 3),黄化が確認される発病評点 2.0 より高い個体を罹病性と判定した場合,抵抗性が 28 個体,罹病性が 86 個体で,1 遺伝子支配の分離比 1:3 にほぼ適合した(表 5).よって,‘ メロン中間母本農 5 号’が有するメロン退緑黄化病抵抗性には,劣性遺伝する 1 つの主要な遺伝子が関与すると推定された.114 個体の F2 を,マーカー遺伝子型(A(‘ メロン中間母本農 5 号’ 型ホモ接合),B(‘AnMP-5’ 型ホモ接合),H(ヘテロ接合))に分類し,それぞれの集団について平均発病評点を求めたところ,A 型集団の平均発病評点は 0.8 となり,B 型集団の 3.2,ならびに H 型集団の 3.1 よりも有意に低い値を示した(表 4).また,発病評点が 0 ~ 1.00 以下であった F2 集団において,18 個体のうち 17 個体のマーカー遺伝子型は A 型であった(図 3).このことから,ECM230 は選抜マーカーとして利用可能であると考えられた.



d 一般特性
一般特性の評価は,2016 および 2018 年度に野菜花き研究部門安濃野菜研究拠点(三重県津市)で実施した.‘メロン中間母本農 5 号’,‘春系 3 号’ および I-10 を用い,植物体および果実特性を調査した.栽培は上記の CCYV の感染が収量および果実品質に与える影響の評価における無接種区と同様に行った.
‘メロン中間母本農 5 号’ の葉は緑色で,葉身長は ‘春系 3 号’ と同程度,主枝の節間長および葉柄長は ‘春系 3 号’ よりも長く,I-10 に比べやや短かった(表 6,図 4).主枝の第 11 ~ 15 節に発生した側枝第 1 節の雌花(両性花)着生率は,2 年の平均が 97.9%であり,I-10 の 11.8%に比べきわめて高かった.
‘メロン中間母本農 5 号’ の 1 果重は 1153 g で,‘春系 3 号’ の 1835g に比べ軽く,I-10 と同程度であった(表 7).成熟日数は ‘春系 3 号’ に比べ 10 日ほど早い 44.5 日で,離層が形成されるためへた離れしやすかった.果形は偏平で,花痕部が 57.0 mm で大きく,果皮に溝が認められた(図 5).果皮は淡緑色であるが,過熟になるとクリーム色に変化した.ネットの密度は極疎,盛り上がりは低かった.果肉は,I-10 が白色であったが,‘ メロン中間母本農 5 号’では淡緑色でやや粘質,食味は劣った.糖度は I-10 より高く,‘ 春系 3 号’より低かった.日持ち性は,I-10 と同程度で,‘ 春系 3 号’より短かった.




2 用途,利用上の注意点
‘メロン中間母本農 5 号’ は,メロン退緑黄化病抵抗性メロン品種育成のための母本として用いることができる.本品種のメロン退緑黄化病抵抗性は劣性に遺伝するため,‘メロン中間母本農 5 号’ を素材として抵抗性を持つ F1 品種を育成するためには,両親への抵抗性付与が必要である.両親の育成には,本系統を 1 回親とした優良固定系統への戻し交雑が望ましく,メロン退緑黄化病抵抗性に連鎖した DNA マーカーを用いることで抵抗性遺伝子を有する個体を効率的に選抜できる.
CCYV は,日本で初めて報告されたウイルスで,現在では海外での発生も数多く報告されている.ウリ科野菜におけるウリ類退緑黄化病に対する抵抗性素材としては,メロン系統 I-10 の報告(Okuda et al. 2013)のみであり,本素材を利用して育成された ‘メロン中間母本農 5 号’ は,メロン退緑黄化病抵抗性品種を育成するための母本としてきわめて有望である.
I-10 においては,CCYV 接種 6 週間後の植物体内でのウイルス蓄積量が罹病性品種に比べ非常に少なく,病徴が認められないことが報告されている(Okuda et al. 2013,).‘メロン中間母本農 5 号’ が有するメロン退緑黄化病に対する抵抗性は,I-10 と同様な抵抗性機構によると考えられる.2015 年度におけるメロン退緑黄化病抵抗性検定試験では,Okuda et al(2013)の報告と同様に,I-10 および ‘メロン中間母本農 5 号’ の植物に病徴は認められなかった.一方,2017 年度の試験では,I-10 および ‘メロン中間母本農 5 号’ の植物に CCYV の感染による非常に弱い病徴が認められた(表 2).2017 年度の播種日は 7 月 7 日であり,2015 年度の試験に比べおよそ 2 か月早く,接種後の栽培時期が 7 ~ 9 月の夏季であったことから,高温条件等により ‘ メロン中間母本農 5 号’ および I-10 では抵抗性が打破され,発病した可能性が考えられる.
I-10 が有するメロン退緑黄化病抵抗性の QTL 解析において 1 つの主要な QTL が検出されている(Kawazu et al. 2018).この QTL 近傍の SSR マーカー ECM230 を用い,‘AnMP-5’ と ‘メロン中間母本農 5 号’ との F2 集団におけるマーカー遺伝子型と発病評点との関連性を調べた結果(表 4,図 3)より,‘メロン中間母本農 5 号’ は I-10 と同一な QTL を有すると考えられる.しかし,マーカー遺伝子型が抵抗性型(‘メロン中間母本農 5 号’ ホモ型)であっても,一部の F2 個体は発病評点が 2.0 より高い罹病性であったことから,ECM230 は抵抗性遺伝子からの距離がやや遠いか,あるいはメロン退緑黄化病抵抗性に係わる 1 つの主要な QTL 以外の遺伝子座が関与する可能性が示唆される.ECM230 を用いて‘ メロン中間母本農 5 号’と罹病性品種・系統との交雑後代から抵抗性個体を選抜する場合,上記のようにマーカー遺伝子型が抵抗性型の個体の中に低率ではあるが罹病性個体も含まれる.よって,抵抗性個体を確実に選抜するためには,メロン退緑黄化病接種試験も併せて行う必要がある.しかし,メロン退緑黄化病抵抗性を判定するためには,接種後 2 か月程度を要し,早期に判定することは難しい.また,抵抗性の判定のためには,CCYV 保毒虫の管理,ならびに検定圃場または施設が必要である.このような問題を解決するためにも抵抗性に連鎖した DNA マーカーによる早期選抜法はきわめて有効で,接種検定に供試する個体数を大幅に減らすことが可能である.今後,DNA マーカーの利用により,‘メロン中間母本農 5 号’ を母本とした実用品種の早期育成が期待される.
メロン中間母本農 5 号’ の育成に関して,中央農業研究センター久保田健嗣博士にはタバココナジラミのウイルス保毒の有無を検定して頂いた.また,野菜花き研究部門技術支援センター安濃業務科の方々,特に堀文明氏,増地智氏および山下大介氏には多大な業務支援を頂いた.ここに記して感謝の意を表する.
すべての著者は開示すべき利益相反はない.