施設ミニトマトにおいて有機 JAS 規格で実施可能な害虫管理プログラム(IPM プログラム)を検討するために,2009 年~ 2013 年および 2015 年~ 2019 年の栽培期間中,茨城県内の有機認証圃場(面積 11 ~ 22 a)において 2 週間に 1 回,植物上の害虫種と個体数を調査した.主要害虫は,コナジラミ類,アブラムシ類,トマトサビダニであった.コナジラミ類は天敵ツヤコバチ類の放飼により著しい被害がでない程度に抑制した.ワタアブラムシには天敵コレマンアブラバチを用いた.2009 年と 2010 年の接種的放飼ではワタアブラムシがハウス全体に広がったため,2011 年以降はバンカー法を実施することで,継続的に低密度に抑制した.チューリップヒゲナガアブラムシは,2010 年以降,天敵チャバラアブラコバチ(試験的使用含む)を用いて抑制した.2016 年のみは定植直後からのチューリップヒゲナガアブラムシの発生により,天敵による密度抑制が不可能となり,栽培を中止したハウスがあった.これは,育苗圃への害虫の持ち込みが原因であったため,翌年以降はこれを防ぐ作業分担とした.トマトサビダニは 2009 ~ 2011 年は被害株率が 20%以上におよんだが,ミルベメクチン乳剤が有機栽培適合資材となり散布できるようになったため,2012 年以降抑制が可能となった.これらの対策により,当初 2,000 kg / 10 a に満たなかった収穫量が,2019 年には 5,000 kg に達した.これらの圃場での実践を経て確立した IPM プログラムを提示した.
有機農業の推進に関する法律(平成 18 年法律第 112 号)で定義される有機農業など,化学合成農薬を削減した栽培においては,病害虫の問題が大きい.例えば,トマトを無農薬で栽培した場合には,減収率は最大で 93%,平均でも 36% と,大きな被害を受けることが報告されている(日本植物防疫協会 1993).葉かび病,疫病などの病気に加え,コナジラミ類,アブラムシ類などの害虫も問題となる.試験圃場での無農薬・有機質肥料でのトマト栽培においても,害虫発生の増加と収量の減少が報告されている(中山ら 1993).有機 JAS 規格で許容される資材を用いたトマト栽培の試験圃場での研究事例では,コナジラミ類への天敵放飼やハダニ類へのマシン油散布などの対応が必要であった(野村ら 2004).
害虫防除の基本は総合的害虫管理(IPM)である.耕種的,物理的,化学的,生物的な手法を,経済性を考慮しつつ矛盾無く組み合わせて用いることにより,害虫密度を許容水準以下に抑制していくものである.有機JAS規格による野菜生産では,化学合成農薬・肥料および遺伝子組換え技術の不使用が必須条件である.このため,化学合成農薬を使っても抑制できない難防除害虫に加え,化学合成農薬を使わないことにより顕在化する害虫も問題となる.しかも,一旦害虫が発生すると,その被害を食い止めることは難しい.従って,有機 JAS 規格に適合する野菜生産では,害虫が発生してからの対処策ではなく,害虫が発生しないよう予防策を中心とした害虫管理プログラム(IPM プログラム)が必要となる.有機農業での害虫管理は,予防的な方法から始めて,対処療法的な方法を組み込んでいく 4 つの段階で考えていく必要がある(Zehnder et al. 2007).第 1 段階は害虫の発生やその被害が起きにくい圃場環境整備である.第 2 段階は,地域の土着天敵類を強化するための植生管理である.第 3 段階は天敵昆虫の放飼である.そして,第4 段階は生物あるいは鉱物由来の殺虫剤の散布などである.
第 1 段階の害虫の問題が起きにくい圃場環境整備は,具体的には,立地や,作付け(時期,輪作,混作),品種(耐性)などの選択による対応である.多くの場合,与えられた立地条件に適した品目を適期に栽培していくことになる.気象条件の変動に対してある程度安定した栽培を志向する場合には,施設栽培も選択肢の一つである.施設栽培の場合には,被覆資材により外部環境と遮断するため大型の害虫類は防げるが,隙間などから侵入する微小な害虫が問題となる.また,こうした施設栽培では,第 2 段階の対応で発生する土着天敵は,圃場への侵入が往々にして害虫よりも遅れるため,第 1 段階の対応の強化(防虫ネットなど)や第 3 段階,第 4 段階まで組み合わせた総合的な管理を考えていく必要がある.一方,露地栽培の場合には,日本では第 3 段階で使える天敵製剤は限られているため(根本 2016),第 1 段階(適地適作,輪作,抵抗性品種など),第 2 段階を中心として第 4 段階を加えた体系を考えることになる.
日本の有機野菜栽培に適用できる害虫防除技術について,科学的な検証を加えた研究としては,試験圃場では赤池ら(2004),塩野(2005),萩森ら(2007)などがあり,また,実際の有機野菜栽培圃場では,長坂ら(2009),赤池ら(2011)などの報告があるが,未だ不十分な状況である.さらに,第 3 段階の天敵活用となると,有機農業では世界的に見ても研究例は少ない(Zehnder et al. 2007).
本研究の目的は,有機 JAS 規格に適合する野菜の施設栽培において,葉菜類を中心とした輪作の中に,春から秋にかけてミニトマトを組み込んだ場合の害虫対策を,経営的に見合う形で体系化することである.このため,先の Zehnder et al.(2007)の第 1 段階として,輪作や太陽熱処理が計画的に実施されているビニールハウスにおいて,開口部を防虫ネットで覆うなどにより害虫の発生しにくい圃場環境を整備した.被覆等により第 2 段階の土着天敵の効果は期待しにくいため,第 3 段階の天敵製剤の利用を防除手段の基幹技術とした.これらの対策で防除できない害虫については,第 4 段階にあたる有機 JAS 規格で使用可能な殺虫剤を用いた.このような組み立てに基づき,実際に有機野菜を出荷する生産法人圃場において,害虫発生の調査とそれに基づく防除対策を 10 年間にわたり積み上げ,生産現場で持続可能な IPM プログラムの構築とその検証を行った.
1.実証圃場と栽培管理
調査は茨城県小美玉市にある株式会社ユニオンファームの有機 JAS 認証圃場で行った.この生産法人では 2001 年の有機 JAS 認定以来,ビニールハウス 150 ~200 棟で主に葉菜類を栽培してきた.輪作を基本とし,夏季には計画的に太陽熱処理を実施した.害虫の発生源となる雑草対策として,ハウス周囲の地面を防草シートで覆った.ミニトマトの栽培は,6 カ所の直営農場のうち,年によって場所を変えながら 3 カ所の農場のハウスで実施した。
ミニトマトの苗は育苗用ビニールハウス(面積約 2 a,栽培棚に電熱線温床を設置して,夜間はトンネル状にビニールで覆い,最低温度を 13℃以上に維持)にて2 月に播種した.第1花房の開花から満開までの苗を定植用に用意した.本圃は面積 2 ~ 3 aの単棟あるいは面積 10~ 22 a の連棟のビニールハウスで(表 1,無加温),おおむね 3 月末に初回定植を行い,作業分散のため複数回に分けて定植し,10 ~ 11 月まで栽培した.ハウス内の畝と通路にはビニールマルチを敷設した.播種,育苗,定植,潅水,施肥,剪枝,葉かき,誘引,収穫などの栽培管理やその作業労力の管理は生産法人が実施した.
ミニトマトの品種(表 1)としては,2009 年と 2010 年は「千果」を利用した.この品種のハウス栽培には,受粉昆虫が必要だったため,セイヨウオオマルハナバチを用いた.しかし,夏の高温により十分働かず,結実が悪かったため,2012 年以降は,単為結果性の品種(「CF ネネ」または「べにすずめ」)に切り替えた.

病害虫防除については,有機 JAS 規格で許容される生物的防除資材あるいは農薬の中から,ミニトマトでの使用基準に基づいて種類,施用量,使用時期,回数を選択し(Appendix 1),防除作業は生産法人が実施した.害虫について大きな問題が生じた場合には,適宜防除方法や防除時期等を変更した.害虫の問題が軽減されるようになると,病害の問題が目立つようになり,2015 年以降は 5 月から病害対策として有機 JAS で使用が認められている殺菌剤の定期的散布を実施した.
2.天敵製剤の放飼
害虫発生時の天敵製剤の放飼は以下の通りである.コナジラミ類の天敵については,6 月以降のコナジラミ類初発確認後,ツヤコバチ類を 1 ~ 2 週間間隔で 9 月まで放飼した.マミー(コナジラミにツヤコバチが寄生して蛹となったもの)のカードは,葉陰に位置するようにミニトマトの葉柄などに吊した.使用量は 25 ~ 30 株あたり 1 カード(10 a あたり 3,000 頭)とした.各年はじめの 2 ~ 3 回は,オンシツツヤコバチ(Encarsia Formosa Gahan)とサバクツヤコバチ(Eretmocerus eremicus Rose and Zolnerowich)の両方,それ以降はサバクツヤコバチのみを放飼した.
アブラムシ類の天敵コレマンアブラバチ(Aphidius colemani Viereck)は,2009 年と 2010 年は本圃でのアブラムシ確認後に規定量を放飼した(10 a あたり 500 ~1,000 頭).2011 年以降は,バンカー法を実施した.バンカー法は害虫の発生前から継続的に天敵を維持するシステムであり,害虫侵入直後の低密度状態から天敵を働かすことができる(根本 2003;長坂ら 2010).天敵を維持するための代替餌(作物の害虫とはならないもの)とそれを維持するための植物(バンカー植物)を設置することにより,天敵銀行(バンカー)として機能する.バンカー植物としてオオムギを 3 月にハウス内に播種し(単棟ハウスでは側窓下に 10 ~ 20 カ所,連棟ハウスでは棟間の柱列に約 10 m 間隔),代替餌としてムギクビレアブラムシ(Rhopalosiphum padi (L.))をオオムギに接種した後,ミニトマトの定植と同時にコレマンアブラバチを放飼した.5 ~ 6 月にはソルゴー(矮性品種)を同様に播種し,トウモロコシアブラムシ(Rhopalosiphum maidis (Fitch))あるいはヒエノアブラムシ(Melanaphis sacchari (Zehntner))を接種した.これらのアブラムシはコレマンアブラバチの寄主となるとともに,土着の寄生者や捕食者の餌にもなる.2010 年以降チューリップヒゲナガアブラムシ(Macrosiphum euphorbiae(Thomas))が発生した場合には,天敵チャバラアブラコバチ(Aphelinus asychis Walker)を規定量(10 a あたり 2,000 頭),1 週間間隔で 3 回程度,アブラムシのコロニーの近くを重点的に,ハウス全体に放飼した.
3.2015 年の無放飼区
2015 年のみは,天敵の導入による害虫抑制効果を検証するために,ツヤコバチ類やアブラバチ類などの天敵製剤を導入しない天敵無放飼区(面積 2.1 a の単棟ビニールハウス)を設けた.微生物製剤等については無放飼区でも天敵放飼区と同様に散布した.ただし,隣接したハウスでは葉菜類が栽培され,アブラムシ対策としてバンカー法が実施されていた.
4.圃場調査
2009 年~ 2013 年と 2015 年~ 2019 年に各種害虫の発生を調査した.2009 年は連棟ハウスにて 5 月から9 月の間,2 週間に 1 回,圃場内からランダムに 26 株~ 46 株を選び,全葉に発生しているアブラムシ類,ハモグリバエ類(潜葉中の幼虫・蛹),ハダニ類(雌成虫数)および天敵アブラバチ類のマミー(アブラムシにアブラバチが寄生して蛹となったもの)を計数した.コナジラミ類については発生の有無のみの確認とし,計数はしなかった.2010 年~ 2013 年は,連棟ハウスあるいは単棟ハウスにて,4 月~ 10 月はじめまで,2 週間に 1 回,合計 13 株~ 130 株について,コナジラミ類(成虫・幼虫)やその寄生蜂のマミーを含め,株上のすべての害虫および天敵類を計数した.微小な害虫・天敵昆虫の計数にはヘッドルーペ(1.3 倍)あるいはルーペ(10 倍)を用いた.トマトサビダニ(Aculops lycopersici Massee))については,主枝が褐色に変化するなど,明確な症状がみられた株の数を適宜調査した.
2014 年は,IPM プログラムが生産者によって実行可能かどうかを明らかにするため,害虫の発生状況の調査は行わず,生産者自身によるモニタリングを基にした害虫防除とした.
2015 年から 2019 年は,連棟ハウスあるいは単棟ハウスにて,4 月~ 10 月の間,2 週間に 1 回,計 60 ~ 210 株について,上,中,下 3 葉と 1 花房で,すべての害虫および天敵類を計数した.各単棟ハウスでは,30 株を上限として調査した.害虫の発生が極端に多い場合には調査株数を減じた.
5.バンカー植物上の天敵の調査
バンカー植物上の天敵類の調査は,2012 年以降,オオムギ,あるいはソルゴーの群落(約 1 m 以内)を 1 カ所のバンカーとして,アブラバチ類のマミー,アブラコバチ類のマミー(アブラバチ類よりも小型で黒色),ショクガタマバエ(Aphidoletes aphidimyza (Rondani)),テントウムシ類,ヒラタアブ類の有無をすべてのバンカー上で記録した.アブラバチ類には放飼したコレマンアブラバチ(マミーは茶色)のほか,ナケルクロアブラバチ(Ephedrus nacheri Quilis,マミーは黒色)など土着のアブラバチ類も見られたので,区別して有無を調べた.
6.統計解析
トマトサビダニ以外の主要な害虫および天敵の個体数データは,ハウスごと調査日ごとに株あたり密度として集計した.年平均密度は調査日ごとに平均した後(複数のハウスがある場合),年ごとの平均値を求めた.主要害虫の密度を年次間で比較するため,ハウスごと調査日ごとの密度を対数変換(log(x+0.5))し,JMP12.2 を用いて統計解析を行った.調査方法が 2009 年~ 2013 年と 2015 年~ 2019 年とで異なるため,両者別個に解析した.コナジラミ類とハモグリバエ類の密度については,定植後日数に対して直線的に増加する傾向が見られたので,一般線型モデルにより年次間で密度増加の程度(定植後日数と年次の交互作用)を分析した.アブラムシ類はおおむね山型の発生消長を示したので,反復測定分散分析により年次間比較を行った.トマトサビダニによる被害株率(総株数に対する被害株数の割合)は,ロジステック回帰分析により,年次間差を解析した.
7.収量データ
実証圃場での収穫量はハウスごと収穫日ごとに記録がなされた.これを年ごとに集計して,総収穫量と 10 a あたり収穫量を算出した.
1. 主要害虫発生の年次変動
実証圃場の有機施設栽培ミニトマトにおける主な害虫は,コナジラミ類,アブラムシ類,トマトサビダニであった(図 1,2).コナジラミ類はオンシツコナジラミ(Trialeurodes vaporariorum Westwood)とタバココナジラミ(Bemisia tabaci (Gennadius))が毎年混発した.アブラムシ類は,チューリップヒゲナガアブラムシとワタアブラムシ(Aphis gossypii Glover)の他,モモアカアブラムシ(Myzus persicae (Sulzer))も見られた.このほか,Liriomyza 属のハモグリバエ類,ヒラズハナアザミウマ(Frankliniella intonsa (Trybom)),ネギアザミウマ(Thrips tabaci Lindeman)などのアザミウマ類,カンザワハダニ(Tetranychus kanzawai Kishida)などのハダニ類,ハスモンヨトウ(Spodoptera litura (Fabricius))などのチョウ目害虫も見られた.
天敵類は,コナジラミ対策として放飼したツヤコバチ類,アブラムシ対策として放飼したアブラコバチ類やアブラバチ類のほか,土着のショクガタマバエも見られた(図 1).

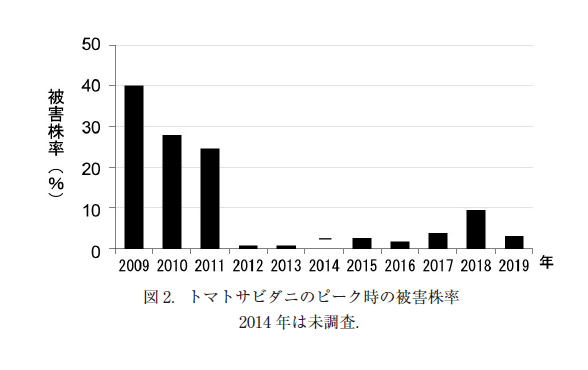
主要害虫の発生量の年次変動(図 3)をみると,コナジラミ類の発生密度は,2017 年のみ,株あたり(3 葉+1 花房)90 頭以上と高い密度となったほかは,おおむね 20 頭/株以下であった.毎年,季節とともに増加傾向(一般線型モデル:前半,後半ともにp<0.0001)にあり,その増加速度については前半 4 年(2010 ~ 2013 年)では年次間差は認められなかった(一般線形モデルの交互作用:df=3, F=2.26, p=0.0693)が,後半 5 年(2015 ~ 2019 年)では年次間差が認められ(一般線形モデルの交互作用:df=4, F=14.36, p<0.0001),特に増加が激しかったのは 2017 年であった.ワタアブラムシは 2009 年と2010 年に多かった(反復測定分散分析:前半 5 年df=4, F=6.45, p=0.0003,後半 5 年df=4, F=1.38, p=0.2518).チューリップヒゲナガアブラムシは2016 年に株あたり(3 葉+ 1花房)100 頭近くにまで増加し,他の年に比べて極端に多かった(反復測定分散分析:前半5 年df=4, F=0.26, p=0.0446,後半 5 年df=4, F=46.39, p<0.0001).トマトサビダニは 2011 年まではピーク時には 20%以上の株で被害が認められたが,2012 年以降は,それ以前より有意に低いレベルに抑えられた(ロジステック回帰分析 df=1, 尤度比カイ二乗=2017.92, p<0.0001,図 2).ハモグリバエ類は 2010 年と 2011 年でやや目立った(一般線形モデルの交互作用:前半 5 年df=4, F=7.97, p<0.0001,後半 5 年df=4, F=2.94, p=0.0214)が,全体的に低いレベルであった(図 3).

2. 主要害虫への対応状況
1) コナジラミ類
コナジラミ類に対しては,成虫あるいは幼虫の発生確認後に,天敵ツヤコバチ類を 1 ~ 2 週間間隔で複数回放飼した.発生の多かった 2017 年は,10 回にわたりオンシツツヤコバチとサバクツヤコバチの両方,あるいはサバクツヤコバチのみの放飼を実施したが,コナジラミ類の増加を止めることができなかった(図 4 左).2017 年の天敵ツヤコバチ類の密度変動を見ると,7 月から8 月にかけて増加しない時期があり,この間にコナジラミ類の密度が大幅に増加した(図 4 左).株あたり 100 頭を越えた 9 月以降は,所々でコナジラミ類の排泄物による果実の汚れ(すす病)が見られた.一方,2018 年は,8 回にわたりオンシツツヤコバチとサバクツヤコバチの両方,あるいはサバクツヤコバチのみの放飼を実施し,9 月に入ってコナジラミ類の増加を止めることができた(図 4 右).2018 年では天敵ツヤコバチ類が連続的に増加することにより,コナジラミ類の増加が止まり,減少に転じた(図 4 右).そして,コナジラミ類の排泄物による果実の汚損はごく一部に限られた.2017 年以外の年は,コナジラミ類,天敵ツヤコバチ類ともに 2018 年と同様の推移を示しており,天敵ツヤコバチの放飼により通常はコナジラミ類をおおむね被害のない程度に抑えられた.なお,2017 年のツヤコバチ類の増加の阻害については,アザミウマ類への対処薬剤による影響が認められた(図 4 左のスピノサド顆粒水和剤).

2) ワタアブラムシ
ワタアブラムシに対しては天敵コレマンアブラバチを放飼した.2010 年は 4 月のワタアブラムシの発生確認後,5 月に 2 回コレマンアブラバチを接種的放飼したが,ワタアブラムシがほぼ観察されなくなったのは 7 月であった(図 5 左).この間,ほぼ全株にワタアブラムシが発生する状態となった後,天敵に寄生されマミーとなった個体が目立つようになり,アブラムシ密度が低下した.この前年の 2009 年は,コレマンアブラバチの接種的放飼の他,気門封鎖剤や糸状菌剤を散布したものの同様の発生推移であった.

2011 年以降は,定植直後から天敵コレマンアブラバチが生息できるように,バンカー法を実施した.バンカー上の天敵類の消長の典型例として 2018 年の調査結果を示した(図 6).4 ~ 5 月には放飼したアブラバチ類が多かったが(図 6 左),6 月には減少し,7 月には土着のアブラバチ類(ナケルクロアブラバチやニホンアブラバチ(Lysiphlebia japonica (Ashmead))など)やアブラコバチ類が増加した.別の年の調査では,アブラコバチ類ではフツウアブラコバチ(Aphelinus varipes (Foerster))が主要種であり,チャバラアブラコバチはほとんど見られなかった(データ省略).8 月にこれらが減少するころには,土着のショクガタマバエが増加した(図 6 右).9 月以降,再放飼されたアブラバチ類も加わり,栽培期間を通して天敵が存在する状態となった.なお,捕食性天敵のヒメカメノコテントウ(Propylea japonica (Thunberg))を放飼した場合には,バンカー上で継続的に観察された(図 6 右)ものの,ミニトマト上ではほとんど見られなかった.

こうしたバンカー法の結果として,栽培初期(5 ~ 6 月)のワタアブラムシの発生はごくわずかとなった(図 5 右).9 月にもワタアブラムシが発生したが,これに対しては,発生部位を切り取ってハウス外に持ち出すことで被害を抑えた.2011 年以降はほぼ同様の対応により,安定してワタアブラムシを低密度に抑えた(図 3).
3) チューリップヒゲナガアブラムシ
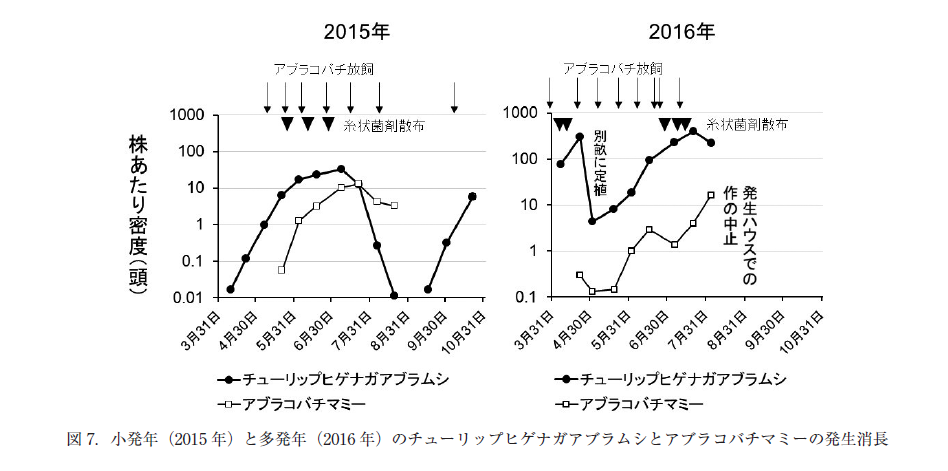
チューリップヒゲナガアブラムシに対しては,発生確認後にまず捕殺した後,チャバラアブラコバチを複数回放飼(2015 年は7 回)した.2016 年以外の年では,この天敵によりチューリップヒゲナガアブラムシの増加を止め,減少させた(図 7 左).発生源は出入り口などからの侵入のほか,前作のレタスで発生した個体が残渣に残っていた場合があった(2015 年).
2016 年のみ,育苗ハウスでチューリップヒゲナガアブラムシの発生が認められ,捕殺と気門封鎖剤の処理を 3 回実施した後に定植したにもかかわらず,本圃において4月はじめから高い密度となった(図 7 右).同じ連棟の別畝に 4 月下旬に定植したミニトマトにも発生が拡大し,その後も密度の高い状態が続いた.天敵チャバラアブラコバチを 8 回放飼し,寄生により生じたマミーが増加したものの,新芽をチューリップヒゲナガアブラムシが覆う状態となったため,この連棟ハウスでは 7 月に栽培を中止した.糸状菌剤も計 5 回散布したが,状態の改善には至らなかった.
なお,梅雨の時期や秋の長雨の時期には,糸状菌殺虫剤(ボーベリアバシアーナ乳剤,ペキロマイセステヌイペス乳剤など)の散布も加え,アブラムシ類とコナジラミ類への対策とした.菌糸の見られる死虫が確認されることもあった.
4) トマトサビダニ,ハダニ類
トマトサビダニに対しては,2011 年までは被害株(茎が茶色に変色し葉が枯れ始めたもの)を見つけ次第切り取って除去したが,発生の拡大は止められず,25%以上の株に被害が及んだ(図 2).2012 年以降は JAS 規格の改正によりミルベメクチン乳剤の使用が 1 作あたり 2 回まで許容されるようになったため,この剤を発生初期に散布することにより被害株率は 10%以下となり,被害株を切り取って持ち出す必要もなくなった.
ハダニ類は局所的に発生し,発生株では下葉から枯れ始めた(データ省略).しかし,被害の拡大は緩やかだったので,大きな問題は生じなかった.
5) ハモグリバエ類,アザミウマ類
ハモグリバエ類については,前半 5 年間(2009 年~2013 年)は潜葉痕が目立ったが,後半 5 年間(2015 年~ 2019 年)は比較的発生が少なかった(図 3).前半 5年はハウス側窓の防虫ネットの目合いが 1 mm であったのに対し,後半 5 年は 0.6 mm だった(表 1).前半 4 年のうち,2010 年と 2011 年は 2012 年と 2013 年に比べて多かった.土着の寄生蜂類の発生も確認しており,季節とともに寄生率の増加が認められた(データ省略).最も発生の多かった 2011 年に天敵イサエアヒメコバチ剤を 1 回放飼した他は,特段の対処は必要とならなかった.
アザミウマ類については,目立って増加することはなかったが,果実に被害が現れることがあり,生産者の判断でスピノサド顆粒水和剤が散布された.例年はアザミウマ類による顕著な被害が認められたハウスのみに散布していたが,2017 年 7 月のみ,全圃場でこの剤が散布された.
3. 天敵放飼による害虫抑制効果の検証(2015 年)
天敵放飼による害虫抑制効果を検証するため,2015 年のみ,天敵を放飼するハウス(放飼区)の他に天敵を放飼しないハウス(無放飼区)を設けた.ただし,無放飼区のハウスに隣接するハウスではバンカー法等により天敵を利用していたため,無放飼区にも天敵類が侵入した.また,コナジラミ類にも土着の寄生蜂類が発生した.こうした攪乱要因はあったものの,結果として,図 8 の通り,コナジラミ類とワタアブラムシ,モモアカアブラムシについては,放飼区の方が無放飼区よりも発生密度が低い状態で推移した(コナジラミ類(一般線型モデル):df=1, F=9.8245, p=0.0045,ワタアブラムシ(反復測定分散分析):df=1, F=5.6235, p=0.0338,モモアカアブラムシ(反復測定分散分析):df=1, F=5.4294, p=0.0366).チューリップヒゲナガアブラムシについては,放飼区において,前作のレタスで発生した個体が残渣に残っていたため,ミニトマトでの発生につながった.特に,栽培前半での発生が目立った.天敵アブラコバチを 6 回放飼したが密度が低下するまでに2 カ月程度かかった(図7左,図 8).栽培期間全体でみると,無放飼区と同程度の発生であり(反復測定分散分析:df=1, F=3.2351, p=0.0953),収量の低下を起こすほどではなかった.
無放飼区では,9 月以降のコナジラミ類の密度が株(3 葉+ 1 花房)あたり 100 頭を越え(図 8),排泄物によりすす病が発生した.果実の汚れにより商品価値が損なわれたため,9 月以降は収穫できる果実はごくわずかとなった.

4. ミニトマト栽培 10 年間の収量の年次変動と害虫発生
調査圃場では,最小 11.1 a(2009 年)から最大 21.6 a(2011 年)の面積でミニトマトを栽培し(表 1),総収穫量は最小で 2,103 kg(2009 年),最大で 8,151 kg(2017 年)であった(図 9).10 a あたりの収穫量(反収)は,当初 3 年ほどは 2,000 kg に達しなかった.この期間は,栽培前半はワタアブラムシ,後半はトマトサビダニによる被害が大きかった.バンカー法によるワタアブラムシ対策,ミルベメクチン乳剤の散布によるトマトサビダニへの対応で,害虫被害が軽減されるようになった 2012 年以降,反収は増加し,2013 年には 3,000 kg を越えるようになった.生産法人のみによる害虫のモニタリングや防除要否の判断をした 2014 年でも 3,000 kg の反収を維持できた.しかし,チューリップヒゲナガアブラムシの発生により 2 つの連棟ハウスのうちの片方で栽培を中断した 2016 年には,10 a あたりの収量が 2,050 kg に減少した.2017 年にはコナジラミ類の多発があったが,株あたり密度(3 葉+ 1 花房)が 100 頭以上に達したのは 9 月の後半以降で栽培期間の終盤であったため,減収にはつながらず,4,000 kg を超える反収となった.また,2018 年には萎凋性病害により栽培を中断した単棟ハウスが 1 棟あったが,反収は 4,000 kg を上回った.2019 年度は病害虫の問題はほぼ無く,反収が 5,000 kg を上回った.

5. 施設有機ミニトマトでの IPM プログラム(害虫部分)の組み立て
収量が比較的安定してきた 2015 年以降の IPM プログラムの概要は表 2 の通りである.前節 2.主要害虫への対応状況を月ごとに整理したものである.ハウス開口部への防虫ネットの展帳や,前作の残渣処理,周辺雑草の除去などは,天敵利用をする上での前提条件である.また,太陽熱処理が 2 年以内に実施された圃場を選択する.育苗圃では,害虫の持ち込みを防ぐため,育苗に関わる作業員が,他の圃場で作業した後に育苗圃へ入らないようにするなど,作業区分や作業の流れを整える.さらに,アブラムシ対策としてのバンカー法を育苗圃でも実施する.
本圃の IPM プログラムでは,コナジラミ類に対し,初発確認後,天敵ツヤコバチを 1 ~ 2 週間間隔で 6 ~ 9 回放飼する.ワタアブラムシ,モモアカアブラムシに対しては,本圃が始まる前からムギ類の播種とムギクビレアブラムシの接種を行い,定植と同時に天敵コレマンアブラバチを放飼するようなバンカー法を実施する.このバンカーを夏場にソルゴーでの土着天敵涵養に繋げるため,5 月にソルゴーを播種,6 月にヒエノアブラムシを接種する.8 月以降にワタアブラムシがミニトマトで発生した場合には,天敵コレマンアブラバチを放飼する.チューリップヒゲナガアブラムシに対しては,5 月以降初発確認後天敵チャバラアブラコバチを 1 週間間隔で 2~ 3 回放飼する.梅雨・秋雨の時期には,これらコナジラミ類,アブラムシ類に対して,糸状菌剤の散布を行う.トマトサビダニに対しては被害を確認後,ミルベメクチン乳剤を散布する.本剤は 2 回までしか使えないので,散布のタイミングに注意する.アザミウマ類に対しては,被害果が目立つ場合に限ってスピノサド顆粒水和剤を散布するが,コナジラミ類の天敵ツヤコバチ類に悪影響があるので,限定的に用いる必要がある.ハスモンヨトウやオオタバコガなどのチョウ目害虫に対しては,発生初期にできる限り捕殺したり持ち出したりした後,BT 剤を散布する.

ビニールハウスでの葉菜類を中心とした野菜の輪作のなかで,4 月定植のミニトマトにおける IPM プログラムを 10 年間の発生害虫の調査を基に検討した.前提として,開口部への防虫ネットの使用,ハウスまわりの防草シートなどにより,害虫の侵入を極力阻止した(第 1 段階).この条件で問題となる害虫について,天敵利用による長期抑制(第 3 段階),有機 JAS 規格で許容される散布剤使用(第 4 段階)の順に防除を組み立てていった.1生産法人での実証試験だが,異なった場所,多様なハウスでの長期にわたる試験であることから,一定の普遍性が担保されていると考えられる。
第 1 段階の強化として,防虫ネットは当初,1.0 mm 目合いを用いていたが,2015 年以降 0.6 mm に切り替えられた.これを境に,ミニトマトではハモグリバエ類の発生がほとんど見られなくなった.ハウスは葉菜類との輪作に用いられており,シュンギクなどでもハモグリバエ類が減少した(未公表データ).目合いが細かいほど害虫の侵入阻止効果が高いことが期待され,害虫を通過させない目合いは,マメハモグリバエでは 0.64 mm,タバココナジラミでは 0.46 mm,ワタアブラムシでは 0.34 mmとされている(Bethke and Paine 1991).害虫侵入阻止の一方で,天敵類の侵入も阻害されることが予想される(長坂ら 2014)が,現状として,バンカーがあればショクガタマバエの発生が見られており,本事例では 0.6 mm 目合いが妥当と考えられる.ただし,0.6 mm 目合いであっても,結果の通りコナジラミ類,アブラムシ類の侵入があり,これらに対して第 3 段階の対策(天敵製剤の放飼)が必要となった.
施設有機ミニトマト栽培において問題となる害虫のうち,栽培前半に重要なものはワタアブラムシやチューリップヒゲナガアブラムシといったアブラムシ類であった.2011 年以降,天敵コレマンアブラバチを用いたバンカー法により,栽培初期のワタアブラムシを抑制できた.このバンカー法の有効性は慣行栽培でも示されており(長坂ら 2010),有機栽培にも今後取り入れていくべき技術である.また,天敵の餌とはなるが害虫とはならないアブラムシ類を,季節を通してバンカー植物上で維持することにより,ショクガタマバエなどの土着天敵を涵養すること(第 2 段階)も可能となり,ワタアブラムシが再発生した場合でも,急速な発生拡大は生じなかった.このショクガタマバエのバンカー法も,この天敵が製剤として販売されていた当時に実用化された有効な技術である(安部 2011).
チューリップヒゲナガアブラムシは,ワタアブラムシ対策のバンカー法を実施している条件でも発生がみられた.これは,天敵コレマンアブラバチがチューリップヒゲナガアブラムシに寄生できないためである(Messing and Rabasse 1995;根本 2004;Nagasaka et al. 2020).バンカー上のショクガタマバエがチューリップヒゲナガアブラムシを捕食する能力を持つ(安部 2011)ものの,チューリップヒゲナガアブラムシが問題となりはじめる 5 ~ 6 月は,ショクガタマバエはバンカー上にはまだ発生していなかった.
チューリップヒゲナガアブラムシに対しては,チャバラアブラコバチが 2011 年に農薬登録され,使用できるようになった.チャバラアブラコバチはアブラムシ類が少発生の時点で発生箇所に集中的に放飼することで,効果を得ることができる(中野 2007; 巽・長坂 2016).実際に,2010 年(試験的使用),2011 年,2015 年,2019 年は,この寄生蜂が有効に働いた.ただし,この天敵については,バンカー法が開発されておらず,実際の圃場でもバンカー上ではほとんど観察されなかった.
2016 年には定植直後からのチューリップヒゲナガアブラムシの蔓延により連棟2棟のうち1棟で栽培中止に追い込まれた.天敵チャバラアブラコバチの放飼や糸状菌殺虫剤の散布を行ったものの,十分な効果は認められなかった.一般に害虫が多くなってから天敵類を投入したのでは,害虫密度の制御は困難であるため,害虫密度が高まらないような事前の工夫が必要となる(矢野 2003).特に育苗ハウスでの害虫の発生は,即効的な化学合成農薬の使用が許されない有機栽培では致命的となる.
2016 年はレタスでチューリップヒゲナガアブラムシが発生し,作業者がその圃場を片付けた後に,灌水のために育苗圃に入ったことが,育苗圃での発生の原因と考えられた.このことから,作業者の動線についても,害虫の持ち込みを防ぐような工夫が必要である.さらには,農場全体で害虫の発生が少ない状態を保つことも必要である(第 1 段階).この生産法人のミニトマト栽培においては,各作業者が担当する作業区分を明確にする対策を導入したことで,2017 年以降問題は生じていない.
コナジラミ類は,5 月末~ 6 月に発生しはじめ,作期の終盤に向かって増加傾向となった.トマトのオンシツコナジラミに対しては天敵ツヤコバチ類の活用で抑制している例がある(小林ら 1996; 佐藤ら 2002; 根本 2016).しかし,タバココナジラミに対しては,サバクツヤコバチを通常の放飼方法で用いても,密度減少に転ずるまでには時間がかかり,年によっては 10 月まで増加傾向のままとなることがあった.「入れない,増やさない,出さない」といったコナジラミ類防除の基本原則(本多 2010)に従っているものの,「増やさない」という部分では有機 JAS 範囲内という限界がある.より安定した生産を行うためには,この問題を解決する必要がある.また,アザミウマ類対策で用いるスピノサド顆粒水和剤は長期間ツヤコバチ類に影響を及ぼす(杉山 2011)ため,2017 年の本圃場のようにツヤコバチ類が増加しない場合(図4 左)もあることにも留意する必要がある.
近年,ナスやピーマン,キュウリ,トマトにおいてタバコカスミカメ(Nesidiocoris tenuis (Reuter))の活用技術が開発されつつある(中石 2014;農研機構 2015;日本・岩本 2020).この天敵は,コナジラミ類やアザミウマ類を捕食する一方で,植物質も餌とする.このため,天敵の密度が増加した場合にはミニトマトでは植物体や果実への被害が懸念され(農研機構 2015),使い方が難しいとされている.今回提示したミニトマトの事例では,前半はツヤコバチ類により密度増加を抑制しつつ,作期の終盤でタバコカスミカメを用いることが考えられる.例えば,タバコカスミカメを 8 月に放飼すれば,この天敵の密度が増加する時期は作期の終了に近い 10 月となり,問題は少ない.タバコカスミカメは現在農薬登録申請中であるが,販売が開始されれば,こうした試みも可能になる.
アブラムシ類やコナジラミ類の増加を抑制する方法として,天敵昆虫の放飼の他にも,糸状菌殺虫剤の散布がある.糸状菌剤は高湿度条件で効果を発揮するため,梅雨や秋雨の時期などに活用し,今回の事例でも,少なくとも補助的な抑制効果はあったものと考えている.この 10 年間の試みでも,天敵昆虫の放飼だけでは害虫の抑制が困難となる場面を経験してきた.糸状菌殺虫剤を有効に使う技術を検討していく必要があるだろう.
以上のように,天敵製剤の利用(第 3 段階)を中心にミニトマトでの IPM プログラムを検討してきたが,ここまでの方法では対処できなかった害虫がトマトサビダニである.2009 年からの 3 年間は,発生株を切り取って持ち出すこととなり,大幅な収量減となった.この害虫は体長 0.2 mm と小さく,防虫ネットで侵入を防ぐことはできない.また,活用できる天敵も今のところ無い.2012 年以降は,ミルベメクチン乳剤の使用が許容されたことにより,トマトサビダニの制御が可能となった.第 1 段階,第 3 段階が有効ではないために,第 4 段階の手段を用いざるを得ない害虫である.
このようにミニトマトの IPM プログラムを組み立ててきたが,広く適用可能かどうかを確認する必要がある.例えば,①生産者から見た技術的な難易度はどうか,②生産者の作業体系に組み込めるものかどうか,③コストと収量維持とのバランスの面で経営的に成り立つかどうか,である.①の生産者からみた技術的な難易度については,2014 年に研究者による害虫調査をせずに,生産者がモニタリングと防除の判断をする栽培を実施することにより検証した.生産者に 5 年間のミニトマト有機栽培の実施経験があるという前提だが,2014 年には大きな問題は生じなかった.経験を積む必要はあるが,生産者に実施可能な技術と判断できる.②の他の作業との関係については,おおむね 2015 年までの積み重ねの中で分担や労力投入の整理がなされて実施可能な状態となっており,また,その後の修正点も加えて,防除の面でも作業効率の面でも効率化された.③の経営的な面については,2015 年~ 2017 年の間の経営的評価(澤田ら 2019)により,法人経営における収支はプラスとなることが報告されている.現地圃場において構築したことで実用的な IPM プログラムとなっている.
IPM プログラムは,これまで慣行栽培において検討がなされてきた.化学合成農薬を使用する慣行栽培では,ミニトマト,トマトともに,コナジラミ類は問題となるが,アブラムシ類は地域によって問題となる程度(根本 2003)で,通常,大きな問題とはならない.また,現状はアブラムシ類に有効な薬剤も利用可能である.しかし,そうした薬剤を使用できない栽培では,ここまで論じてきたようにアブラムシ類が大きな問題となる.すなわち,慣行栽培での害虫発生を基準として,有機栽培の IPM プログラムを考えることは出来ない.一方,慣行栽培においても,天敵等により農薬を削減した場合には潜在的な害虫が顕在化する(中野 2007;長坂ら 2010;柿元ら 2015).トマトサビダニもそうした害虫の一つであり(河合 2003),天敵オンシツツヤコバチを利用して農薬を削減した圃場で認識される事例もある(原田・中野 1999).慣行栽培においても,農薬を使用しない状態から防除を考えていくような IPM プログラムの組み立て方が重要だろう(長坂・杜 2016).
この報告では,害虫対策についてのみ検討してきたが,実際には,病害の対策も組み合わせた IPM プログラムの結果として,収量の増加の確認と経営的評価が行われている(澤田ら 2019).害虫の問題が解決し始めた 2015 年頃から病害も目立ち始めたため,病害対策も検討してきた.これについては別途報告がなされる予定である.
現実の野菜生産では,様々な制約の中で IPM プログラムを組み立てていく必要がある.圃場規模,管理作業,労力配分,品質や収量への厳しさ,などの点から,現実の農場経営において防除コストに見合った増収が確認された事例は希少である.1地域での実証試験ではあるが,10 年間の蓄積から導いた IPM プログラムは,実用的な技術として記録する価値があると考える.




本研究の圃場調査にあたっては,奥圭子博士,勾坂晶博士らのご協力をいただき,調査結果の統計解析には光永貴之博士のご指導をいただいた.また,株式会社ユニオンファームの玉造洋祐社長はじめ従業員の皆様には多大なるご協力と便宜をいただいた.2015 年度の試験の実施に当たっては,平成 26 年度補正予算 農林水産業の革新的技術緊急展開事業(ID14649773)「大規模施設有機野菜生産法人におけるバンカー法等天敵活用による生産安定化の実証」による支援を受け,根本久博士に外部アドバイザーとして数々のご助言をいただいた.これら大勢の皆様に深く感謝申し上げる.
すべての著者は,開示すべき利益相反はない.