原著論文
ジアミド剤散布による2 種カキ害虫フタモンマダラメイガおよびカキノヘタムシガの同時防除
2019 年 2019 巻 2 号 p. 11-20
詳細
2019 年 2019 巻 2 号 p. 11-20
カキノヘタムシガの防除適期に散布するジアミド剤のフタモンマダラメイガに対する同時防除効果を調査した.カキノヘタムシガ第 1,2 世代幼虫の防除適期におけるジアミド剤の散布は,本害虫による果実被害を対照のフェニトロチオン水和剤と同等に抑えることができた.カキノヘタムシガ第 1 世代幼虫の防除適期に散布したフルベンジアミド水和剤のフタモンマダラメイガに対する同時防除効果は,無防除区においてもフタモンマダラメイガ第 2 世代幼虫の密度が低く推移することからその有効性を評価できなかった.その一方で,カキノヘタムシガ第 2 世代幼虫に対する 2 種ジアミド剤の散布は,フタモンマダラメイガ越冬世代(第 3 世代)幼虫の密度抑制に有効であった.フタモンマダラメイガの越冬世代幼虫は密度が高いために防除の必要性が高いが,第 1,2 世代幼虫は密度が低いことから防除の必要はないと考えられた.以上から,フタモンマダラメイガの被害抑制は,カキノヘタムシガ第2 世代幼虫防除時期のジアミド剤散布で可能と考えられた.
フタモンマダラメイガEuzophera batangensis Caradja(チョウ目:メイガ科)は,ヒメコスカシバSynanthedon tenuis(Butler)(チョウ目:スカシバガ科)と同様にカキノキDiospyros kaki Thunb.(ツツジ目:カキノキ科)の枝と幹の分岐部や枝の基部などを食害し,枝の枯死や樹勢低下を引き起こす枝幹害虫である(藤田,林 2015,杖田ら 2014).本種はカキ害虫として知られてきたが,近年はナシでも被害が認められるようになった(笈田ら 2015,下元 2013).本種に対して近年,ジアミド剤を利用した防除の研究が進んだが(新井ら 2019,藤田,林 2015,下元 2013,杖田ら 2014),フルベンジアミド剤の高濃度散布時期である開花期前(藤田,林 2015,杖田ら 2014)や新井ら(2019)が報告した本種のフルベンジアミド剤による防除適期(7 月上旬,8 月下旬)は,カキ栽培における最重要チョウ目害虫であるカキノヘタムシガStathmopoda masinissa Meyrick(チョウ目:ニセマイコガ科)の防除適期(6 月上旬,7 月下旬~ 8 月上旬)と異なることから,フタモンマダラメイガに対する防除により殺虫剤散布回数や防除経費が増加することが懸念される.藤田,林(2015)は,カキノヘタムシガの防除時期と考えられる 6 月上旬または 8 月上旬のジアミド剤の散布により,フタモンマダラメイガの食入幼虫数が少なくなることを報告した.これらのことから,カキノヘタムシガの第 1,2 世代幼虫の防除適期におけるジアミド剤の散布で,フタモンマダラメイガの第 2 世代および第 3 世代(越冬世代)幼虫を同時防除できると考えられる.そこで今回,カキノヘタムシガの防除適期にフタモンマダラメイガにも適用のあるジアミド剤(フルベンジアミド水和剤およびクロラントラニリプロール水和剤)を散布し,フタモンマダラメイガに対する同時防除効果を調査した.
1.調査圃場
調査は 2016 年~ 2018 年にかけて農研機構果樹茶業研究部門ブドウ・カキ研究拠点内のカキ 2 園(A 園と B 園)で実施した.B 園は A 園の約 50 m 北に位置していた.A 園は列間・樹間とも約 5 m,栽植本数は 40 樹,面積は約 12 a,‘早秋’が園の北側に 15 樹,‘ 富有’が園の南側に 14 樹栽植され,他にも複数の品種が混植されていた.A 園における枝幹虫のほとんどはフタモンマダラメイガであった.B 園は列間・樹間とも約 4 m,栽植本数は 11 樹,面積は約 4 a,‘富有’が 10 樹と‘ 富有’に‘ 平核無’を高接ぎした樹が 1 樹栽植されていた.A 園では 2012 年 8 月 9 日に‘ 富有’全樹に,また‘ 早秋’の一部樹に新井ら(2019)が試験散布を行った以外,試験薬剤散布前年まで殺虫剤無散布とした.A 園の‘ 早秋’ではカキノヘタムシガ第 1,2 世代幼虫それぞれの防除適期である「‘ 富有’の満開 10 日後」(新井ら 2016)または「第 1 世代成虫の初発日の 10 日後」(新井ら 2018)に試験薬剤を散布した(Table 1).ただし,2016 年のカキノヘタムシガ第 1 世代幼虫防除適期には殺虫剤無散布とした.A 園の‘ 富有’ではカキノヘタムシガの密度を抑えるため,‘ 早秋’の防除日にあわせて殺虫剤を散布した.B 園では 2012 年 6 月 13 日に全樹に殺虫剤が散布されて以降殺虫剤無散布とし,カキノヘタムシガに対する無防除圃場とした.
2. カキノヘタムシガ幼虫に対するジアミド剤の散布がフタモンマダラメイガ食入幼虫数に及ぼす影響
1)カキノヘタムシガ被害果調査
2016 年~ 2018 年にかけて,A 園の‘ 早秋’結実樹および‘ 富有’全樹,および B 園の‘ 富有’全樹について,地上から2m 以下の高さに結実した全果実についてカキノヘタムシガ第 1,2 世代幼虫による被害を調査した.調査は 2016 年 7 月 14 日と 8 月 22 日,2017 年 7 月 21 日と 8 月 24 日および 2018 年 7 月 17 ~ 18 日と 8 月 21 日に実施した.なお,A 園と B 園とも試験開始前にカキノヘタムシガに対する防除を実施していなかったことや,殺虫剤無散布条件とした期間が異なることから,両園におけるカキノヘタムシガの発生量を事前に把握し,翌年以降の防除試験効果を評価するために,両園の‘ 富有’全樹について,地上から 2 m 以下の高さに結実した全果実について,A 園では 2015 年 8 月 27 日に,B 園では 9 月 10 日にカキノヘタムシガによる被害を調査した.
2 ) カキノヘタムシガ第 2 世代幼虫防除適期散布のフタモンマダラメイガ越冬世代幼虫に対する同時防除効果
フルベンジアミド水和剤のカキノヘタムシガ第 2 世代幼虫の防除適期における散布が,フタモンマダラメイガ越冬世代食入幼虫数に及ぼす影響を 2016 年に調査した.A 園の‘ 早秋’ 13 樹のうち 4 樹を調査樹に,9 樹を対照樹とした.調査樹および対照樹とも,カキノヘタムシガ第 1 世代幼虫防除時期は殺虫剤無散布とし,調査樹では第 2 世代幼虫の防除適期となる 7 月 26 日にフルベンジアミド水和剤(商品名:フェニックスフロアブル)4,000 倍液を,対照樹には同日フェニトロチオン水和剤(商品名:スミチオン水和剤 40)1,000 倍液を幹および枝に薬液が到達するように樹全体に散布した(1 樹あたり約 10 リットル)(Table 1).調査樹および対照樹の全樹について,地上から約 1 ~ 1.7 m の枝幹部に食入した幼虫を 4 月 18 日,7 月 1 日,7 月 12 日,10 月 4 日と 10 月 27 日に粗皮削り用器具を用いて掘り出した.
3) カキノヘタムシガ防除適期 2 回散布によるフタモンマダラメイガ第 2 世代幼虫および越冬世代幼虫に対する同時防除効果
フルベンジアミド水和剤のカキノヘタムシガ第 1 世代幼虫,および第 1,2 世代幼虫防除適期の散布がフタモンマダラメイガ食入幼虫数に及ぼす影響を 2017 年に調査した.A 園の‘ 早秋’ 13 樹のうち 9 樹を調査樹に,4 樹を対照樹とした.調査樹のうち 5 樹はフルベンジアミド 2 回散布区(試験区 1)とし,カキノヘタムシガ第 1 世代幼虫の防除適期である 6 月 6 日と第 2 世代幼虫の防除適期となる 7 月 28 日にフルベンジアミド水和剤 4,000 倍液を散布した.調査樹の 4 樹はフルベンジアミド水和剤 1 回散布区(試験区2)とし,6 月 6 日にフェニトロチオン水和剤 1,000 倍液を,また 7 月 28 日にフルベンジアミド水和剤 4,000 倍液を散布した.対照樹には 6 月 6日および 7 月 28 日にフェニトロチオン水和剤 1,000 倍液を散布した(Table 1).いずれの薬剤も幹および枝に薬液が到達するように樹全体に散布した(1 樹あたり約 10 リットル).調査樹および対照樹の全樹について,地上から約 1 ~ 1.7 m の枝幹部に食入した幼虫を 5 月 31 日,7 月 24 日,10 月 4 日および 10 月 25 日に掘り出した.
4) 2 種ジアミド剤の防除効果比較試験
フルベンジアミド水和剤とクロラントラニリプロール水和剤(商品名:サムコルフロアブル)のカキノヘタムシガ第 2 世代幼虫防除適期の散布がフタモンマダラメイガ越冬世代食入幼虫数に及ぼす影響を 2018 年に比較した.A 園の‘ 早秋’ 13 樹のうち 8 樹を調査樹に,5 樹を対照樹とした.調査樹のうち 4 樹はクロラントラニリプロール水和剤散布区(試験区1)とし,カキノヘタムシガ第 1 世代幼虫の防除適期となる 6 月 7 日にフェニトロチオン水和剤 1,000 倍液を,カキノヘタムシガ第 2 世代幼虫の防除適期となる 7 月 25 日にクロラントラニリプロール水和剤 5,000 倍液を散布した.調査樹のうち 4 樹はフルベンジアミド水和剤散布区(試験区2)とし,6 月 7 日にフェニトロチオン水和剤 1,000 倍液を,7 月 25 日にフルベンジアミド水和剤 4,000 倍液を散布した.対照樹には 6 月 7 日と 7 月 25 日にフェニトロチオン水和剤 1,000 倍液を散布した(Table 1).いずれの薬剤も幹および枝に薬液が到達するように樹全体に散布した(1 樹あたり約 10 リットル).調査樹および対照樹の全樹について,地上から約1 ~ 1.7 m の枝幹部に食入した幼虫を6 月 13 日,7 月 23 日,10 月 2 日および 10 月 30 日に掘り出した.
3. 調査圃場におけるフタモンマダラメイガ発生消長および寄生蜂寄生率の推移
調査圃場におけるフタモンマダラメイガの発生量および発生時期を明らかにするため,A 園の‘ 富有’ 14 樹を調査樹として,調査樹の全樹について,2016 年 5 月 23 日~ 10 月 27 日,2017 年 3 月 14 日~ 10 月 25 日,2018 年 6 月 13 日~ 10 月 30 日の期間に約 2 週間間隔で,地上から高さ約 1 ~ 1.7 m の枝幹部に認められた全ての虫糞について周辺部位の粗皮を除去し,フタモンマダラメイガ食入幼虫を採集した.採集した幼虫は直径 9 cm,深さ 2 cm のプラスチックシャーレ内で,人工飼料(インセクタLFS,日本農産工業株式会社製)を与え,ブドウ・カキ研究拠点内に設置した百葉箱中の自然温度条件で飼育し,羽化したフタモンマダラメイガ成虫および寄生蜂の種を確認した.また,次式から寄生蜂による寄生率を算出した.
寄生率 = 羽化した寄生蜂数 × 100 /(羽化した寄生蜂数+フタモンマダラメイガ成虫数)
なお,‘ 早秋’から採集したフタモンマダラメイガ幼虫の寄生蜂による寄生率も同様の方法で調査した.
4.統計処理
得られた結果の統計解析には「R」Version3.5.2(R Development Core Team 2018)を用いた.カキノヘタムシガの被害果率は Bonferroni の補正を施した有意水準を用い,Fisher の正確確率検定により 2 区間ずつ区間差を比較した.フタモンマダラメイガの枝幹部への食入幼虫数は t- 検定または Tukey-Kramer 法により 5%水準で試験区間の差を比較した.
1. カキノヘタムシガ幼虫に対するジアミド剤の散布がフタモンマダラメイガ食入幼虫数に及ぼす影響
1)カキノヘタムシガによる被害果率
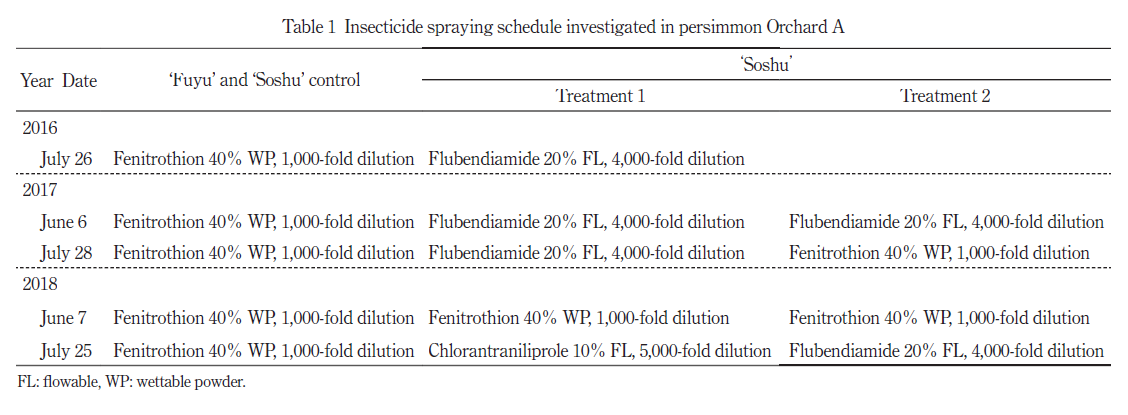
‘ 早秋’におけるカキノヘタムシガ被害果率をFig. 1 に示した.殺虫剤無散布とした 2016 年のカキノヘタムシガ第 1 世代幼虫による対照樹と試験樹の被害果率はそれぞれ 42.4%,50.9% と高かったが,薬剤防除実施後の被害果率は世代を経るごとに低くなる傾向であった.2017 年以降カキノヘタムシガに対する被害果率は試験区間で差は認められなかった.
‘ 富有’における被害果率を Fig. 2 に示した.殺虫剤無散布条件であった 2015 年の第 2 世代および 2016 年の第 1 世代のカキノヘタムシガによる被害果率はそれぞれ 37.5 ~ 47.2%,18.2 ~ 28.0%と高い水準であった.殺虫剤無散布を続けた B 園ではカキノヘタムシガによる被害果率はその後も高く推移した.防除を実施した A 園では 2016 年のカキノヘタムシガ第 2 世代以降,被害果率は B 園と比べて有意に低くなり,‘ 早秋’と同様に年を経るごとに低くなる傾向であった.

2) カキノヘタムシガ第 2 世代幼虫防除適期散布のフタモンマダラメイガ越冬世代幼虫に対する同時防除効果
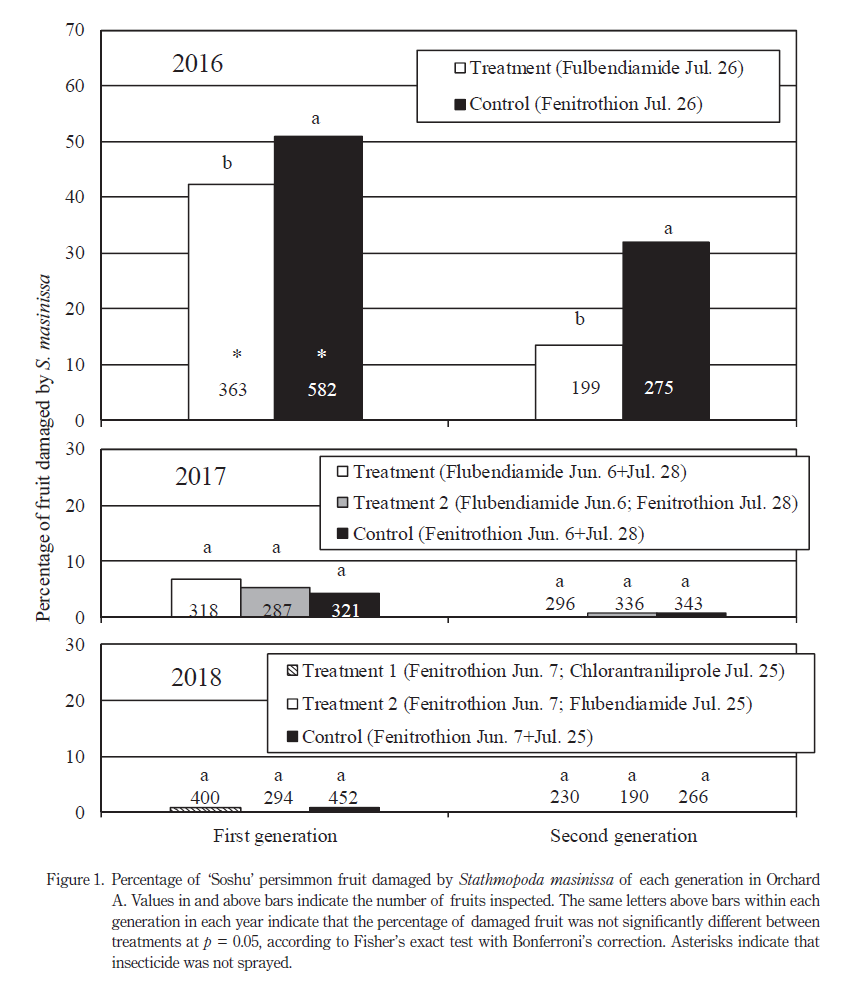
フルベンジアミド水和剤を散布した試験樹および対照樹におけるフタモンマダラメイガの樹あたり食入幼虫数の推移を Fig. 3 に示した.フタモンマダラメイガ食入幼虫数は 7 月までは区間で差は認められなかったが,10 月のフタモンマダラメイガ越冬世代の食入幼虫数は試験樹では対照樹に比べて有意に少なくなった.
3) カキノヘタムシガ防除適期 2 回散布によるフタモンマダラメイガ第 2 世代幼虫および越冬世代幼虫に対する同時防除効果
フルベンジアミド水和剤を散布した試験樹および対照樹におけるフタモンマダラメイガの樹あたり食入幼虫数の推移を Fig. 4 に示した.フタモンマダラメイガ食入幼虫数は,7 月の第 2 世代までは区間で有意差は認められなかったが,10 月の越冬世代では,フルベンジアミド水和剤を 6 月と 7 月下旬の 2 回散布した試験区 1 では対照樹よりも有意に少なく,フルベンジアミド水和剤を 6 月に 1 回散布した試験区 2 では試験区 1 および対照樹と差は認められなかった.

4)2 種ジアミド剤の防除効果比較試験
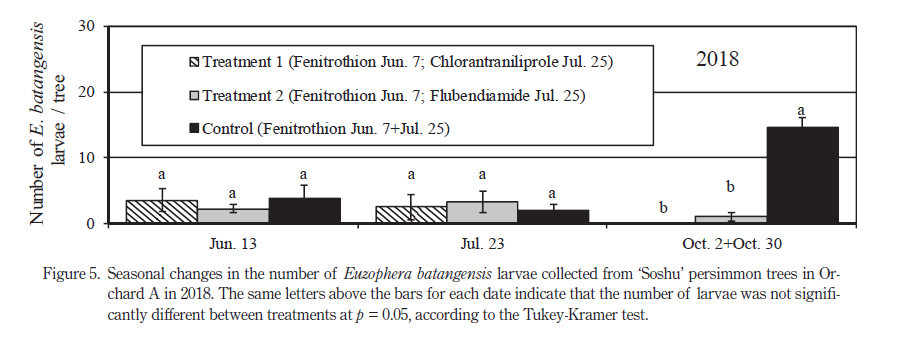
フルベンジアミド水和剤またはクロラントラニリプロール水和剤を散布した試験樹および対照樹におけるフタモンマダラメイガの樹あたり食入幼虫数の推移を Fig.5 に示した.6 月および 7 月のフタモンマダラメイガ食入幼虫数は区間で差は認められなかった.10 月のフタモンマダラメイガ越冬世代の食入幼虫数はクロラントラニリプロール剤散布区とフルベンジアミド水和剤散布区で差は認められず,またいずれのジアミド剤散布樹も対照樹に比べて有意に少なかった.
2. 調査圃場におけるフタモンマダラメイガ発生消長および寄生蜂寄生率の推移
フタモンマダラメイガ食入幼虫数の推移を Fig. 6 に示した.食入幼虫は調査期間を通じて認められたが,いずれの調査年でも 5 月~ 8 月上旬までの第 1,2 世代では比較的少なく,8 月下旬以降の秋季に増加する傾向があった.

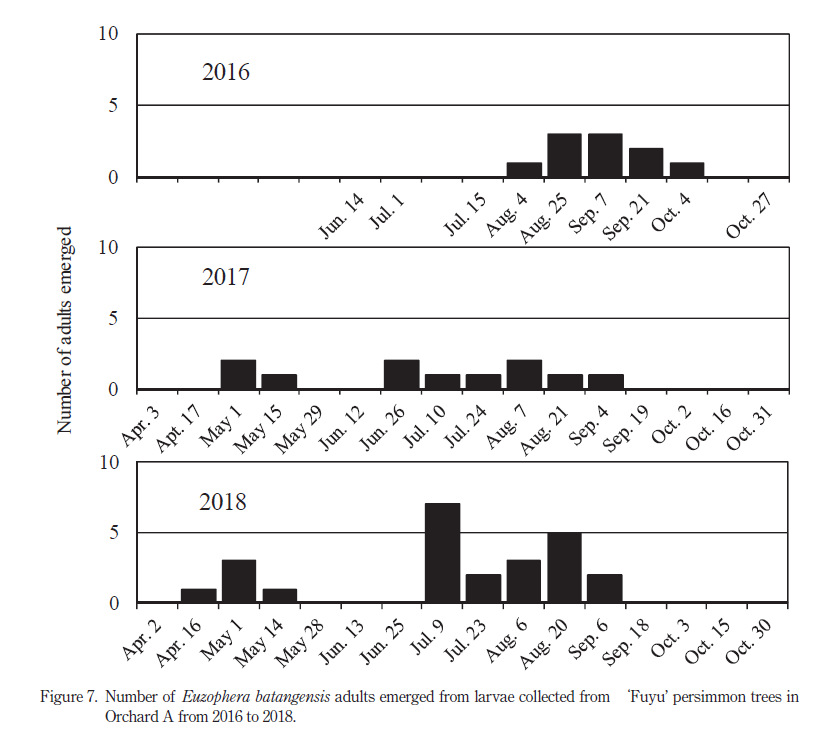
樹皮下から採集したフタモンマダラメイガ幼虫を飼育した結果,本種の越冬世代成虫は 4 月中旬~ 5 月中旬に羽化した(Fig. 7,2017 年,2018 年).本種成虫の羽化は 6 月下旬~ 9 月上旬まで続き,第 1 世代成虫は 6 月下旬~ 7 月上旬頃に,第 2 世代成虫は 8 月下旬~ 9 月上旬に多くなる傾向であった.8 月までに採集したフタモンマダラメイガ幼虫は年内に羽化したが,9 月以降採集された幼虫は翌年羽化する傾向であった.2017 年と 2018 年には 9 月中旬以降に成虫は羽化しなかった.
フタモンマダラメイガ幼虫にはコマユバチ科の単寄生性・内部捕食寄生性の 2 種,コウラコマユバチ亜科コウラコマユバチの一種 Phanerotoma sp. とサムライコマユバチ亜科サムライコマユバチの一種(属未同定)の寄生が認められた.特にコウラコマユバチの一種は,2016 年は羽化した寄生蜂総数の 56.7%,2017 年は 90.7%,2018 年は 86.7%を占めた.寄生蜂による寄生率は,7 月下旬以降に 33 ~ 100%と比較的高く推移した(Fig. 6).ジアミド剤を散布した‘ 早秋’ 調査樹のフタモンマダラメイガの食入幼虫数が少なかったことから,‘ 早秋’ 試験樹および対照樹全樹の食入幼虫の寄生率を算出したところ,寄生率は調査期間中,50 ~ 100%と‘ 富有’と同様に高く推移した(Table 2)

今回,カキノヘタムシガに対する防除で‘ 富有’ における被害果率を殺虫剤無散布園と比べて大幅に低くすることができたことから,カキノヘタムシガの防除時期は適切であったと考えられた.ジアミド剤のフルベンジアミド水和剤またはクロラントラニリプロール水和剤をカキノヘタムシガ第 2 世代幼虫の防除適期である 7 月下旬に散布した樹では,同時期に対照薬剤としてフェニトロチオン水和剤を散布した対照樹と比べてフタモンマダラメイガ越冬世代の食入幼虫数が少なく,さらにカキノヘタムシガ第 2 世代幼虫による被害果率も対照樹と同等に低かった.このことから,カキノヘタムシガ第 2 世代幼虫に対するこれらジアミド剤の散布によりフタモンマダラメイガ越冬世代幼虫の同時防除が可能と考えられた.
フルベンジアミド水和剤の 6 月上旬の防除はカキノヘタムシガ第 1 世代幼虫による果実被害を対照薬剤とほぼ同等に低く抑えることができたが,7 月までのフタモンマダラメイガ幼虫数はフルベンジアミド水和剤散布樹と対照樹で差が認められなかった.‘ 富有’におけるフタモンマダラメイガ幼虫の消長を見ると,フタモンマダラメイガ食入幼虫数は第1,2 世代である 5 ~ 8 月上旬までは少なく,8 月下旬以降多くなる傾向であった(Fig. 6).このことから,6 月上旬の防除効果を検出できなかったのは第 2 世代幼虫の食入幼虫が少ないことが影響したと考えられた.また,今回フルベンジアミド水和剤散布を6 月上旬の 1 回のみとした区では,6 月上旬のフルベンジアミド水和剤のフタモンマダラメイガ越冬世代幼虫に対する効果が判然としなかった.藤田,林(2015)の試験では 6 月 10 日に散布したジアミド剤は越冬世代幼虫の密度抑制効果が劣ることが,また新井ら(2019)の試験では 7 月上旬の本剤の散布は越冬世代幼虫の密度抑制効果が劣ることが報告されている.これら結果に加え,フタモンマダラメイガの第 2 世代は密度が低いことに鑑みれば(Fig. 6),カキノヘタムシガ第 1 世代幼虫に対するジアミド剤による防除によってフタモンマダラメイガ越冬世代幼虫までの同時防除効果を期待するのは難しいと考えられた.
樹皮下から採集したフタモンマダラメイガ幼虫の人工飼料による飼育により判明した成虫の羽化時期は,新井ら(2019)のフェロモントラップでのモニタリング結果と同様の傾向であった.2017 年と 2018 年の幼虫採集と飼育の結果から,本調査地におけるフタモンマダラメイガの生活環は以下のように推測された.越冬世代の羽化は 5 月上旬に多く認められ,その 4 ~ 6 週間後の 5 月下旬~ 6 月中旬に多数採集された幼虫は第 1 世代と考えられた.第 1 世代成虫の羽化は 6 月下旬~ 7 月上旬に多く認められ,その 4 週間後の 7 月下旬~ 8 月上旬には第2世代とみられる幼虫が多く採集された.第 2 世代成虫は 8 月下旬~ 9 月上旬に多く羽化し,その 4 週間後となる 9 月下旬~ 10 月下旬およびそれ以降に採集された幼虫は第 3 世代と考えられた.今回の調査では,9 月以降に採集された幼虫は概ね翌春に羽化する傾向であったことから,本調査地では第 3 世代幼虫での越冬が主体と考えられた.今回得られた成果がフタモンマダラメイガの発生が年 3 回以外の地域でも適用できるのかどうか,今後調査する必要がある.
新井ら(2019)はフタモンマダラメイガの寄生蜂寄生率が高いことを報告したが,今回の調査でも,採集したフタモンマダラメイガから新井ら(2019)が報告した 2 種寄生蜂の羽化が認められ,調査期間中を通じて高い寄生率で推移した.このことから,本種は寄生蜂による密度抑制効果が高いと考えられた.ただし,今回の試験では 9 月以降にフタモンマダラメイガの密度が高くなったが,同時期には天敵による寄生率も高く推移したことから,秋季におけるフタモンマダラメイガの増加は殺虫剤が天敵類に影響したことによる誘導多発生であるとは考えにくい.今後,9 月以降本種密度が高くなる要因について解明する必要があると考えられた.
すべての著者は開示すべき利益相反はない.